Saskaņā ar flagellas atrašanās vietu baktērijas tiek sadalītas. Kustības organelli: funkcijas un struktūra, vienšūņu kustības iezīmes
Struktūra. Apmēram pusei zināmo baktēriju sugu uz virsmas ir kustību orgāni – viļņveidīgi izliekti flagellas. Ziedu masa ir līdz 2% no baktērijas sausās masas. Kaņģa garums ir garāks par mikroorganisma ķermeņa garumu un ir 3–12 μm; flagellum biezums ir 0,02 μm, un polārie flagellas ir biezākas par peritrihiem.
Flagellas sastāv no flagellīna proteīna (latīņu flagella - flagellum), kas pēc savas struktūras pieder pie miozīna tipa saraušanās proteīniem. Karogs satur vai nu vienu viendabīgu proteīna pavedienu, vai 2–3 pavedienus, kas cieši salocīti bizē. Kaņģa vītne ir stingra spirāle, kas savīta pretēji pulksteņrādītāja virzienam; spirāles solis ir raksturīgs katrai baktēriju sugai.
Ziedu skaits, izmērs un izvietojums ir pazīmes, kas noteiktai sugai ir nemainīgas un tiek ņemtas vērā taksonomijā. Tomēr dažas baktērijas var veidot flagellas. dažādi veidi. Turklāt flagellas klātbūtne ir atkarīga no vides apstākļiem: ilgstošas kultivēšanas laikā uz cietas barotnes baktērijas var zaudēt flagellas, un uz šķidrām barotnēm tās var iegūt atkārtoti. Tās pašas sugas karogu skaitu un atrašanās vietu var noteikt pēc dzīves cikla stadijas. Tāpēc nevajadzētu pārvērtēt šīs pazīmes taksonomisko nozīmi.
Baktēriju klasifikācija pēc flagellas skaita un atrašanās vietas:
1. Atriči - flagellas nav.
2. Monotrichus- viens flagellum, kas atrodas vienā no šūnas poliem (Vibrio ģints)- monopolārs monotrichous izkārtojums flagellas, viskustīgākās baktērijas.
3. Polytrichus — daudzi flagellas:
– lophotrichous- flagellas saišķis vienā šūnas polā (dzimšana Pseidomonas, Burkholderia) - monopolārais daudzveidīgs flagellu izvietojums;
– amfitrichous- pie katra šūnas pola atrodas kažogu saišķis (Spirillum ģints)- bipolāri daudzveidīgs flagellas izvietojums;
– peritrichous- flagellas nav izvietotas noteiktā secībā pa visu šūnas virsmu (fam. Enterobacteriaceae(dzimšana Escherichia, Proteus), ģimene. Bacillaceae,ģimene Clostidiaceae), flagellu skaits svārstās no 6 līdz 1000 vienā šūnā atkarībā no baktēriju veida (7. att.).
7. att. Iespējas flagellas atrašanās vietai baktērijās:
1 - monotrich, 2 - lophotrich;
3 - amfitrichous; 4 - peritrihs.
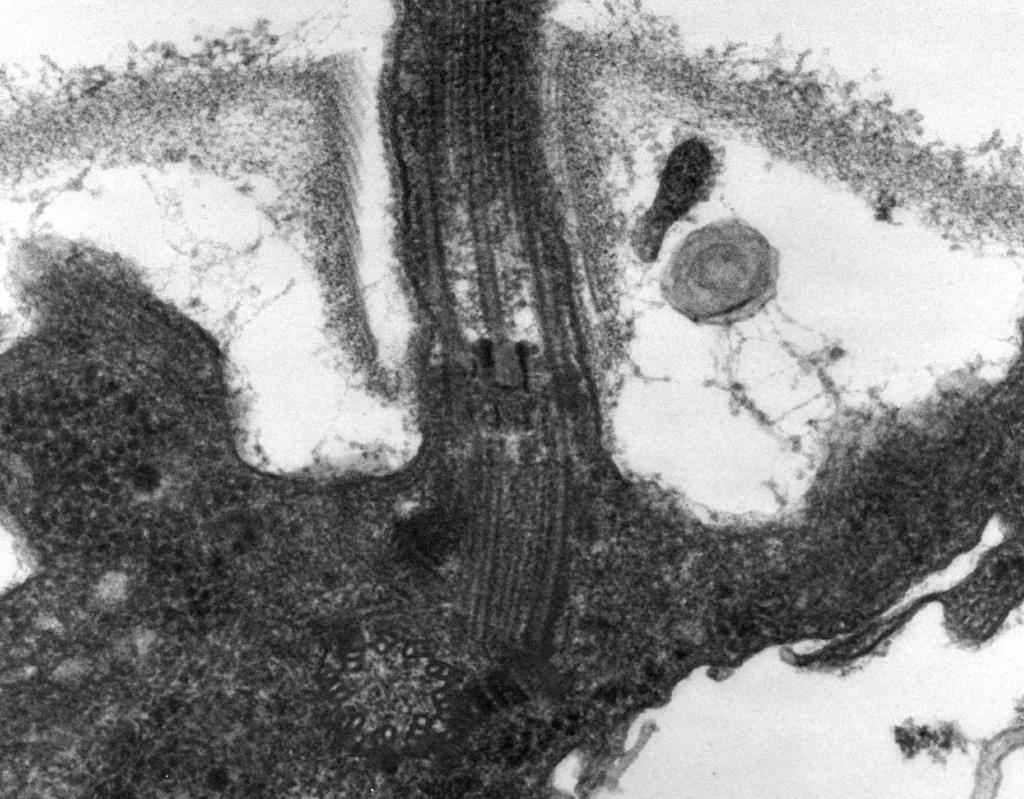
– Elektronu mikroskopija to atklāj flagellum sastāv no trim daļām: spirālveida pavediens, āķis un pamatkorpuss (8. att.).
Galvenā flagellum daļa ir gara spirālveida vītne (fibrila) - stingrs dobs cilindrs ar diametru aptuveni 120 nm, kas sastāv no flagellīna proteīna. Olbaltumvielu molekulas veido 11 rindas visā pavediena garumā un ir sakārtotas spirālē. Vītnes augšanas laikā šūnā sintezētās olbaltumvielu molekulas iziet cauri cilindra dobumam un tās galā tiek piestiprinātas pie spirāles. Kaņģa galā ir proteīna vāciņš (vāciņš), kas aizver cilindra atvērumu un novērš olbaltumvielu molekulu izdalīšanos vidē. Karogas pavediena garums var sasniegt vairākus mikrometrus. Dažām baktēriju sugām karogs no ārpuses ir papildus pārklāts ar apvalku. CS virsmā spirālveida vītne pāriet sabiezinātā izliektā struktūrā - āķī.

Rīsi. 8. Kaņģa struktūras shēma
2. Āķis(20–45 nm biezs) netālu no šūnas virsmas ir salīdzinoši īss cilindrs, kas sastāv no proteīna, kas atšķiras no flagellīna, un kalpo, lai nodrošinātu elastīgu kvēldiega savienojumu ar bazālo ķermeni.
3. Bāzes ķermenis atrodas pie karogdziedzera pamatnes un nodrošina tā rotāciju. Bāzes korpuss satur 9–12 dažādus proteīnus un sastāv no diviem vai četriem diskiem (gredzeniem), kas savērti uz stieņa, kas ir āķa turpinājums. Šie gredzeni ir uzstādīti CPM un CS. Divi iekšējie gredzeni (M un S) ir obligātas pamata korpusa sastāvdaļas. M-gredzens ir lokalizēts CPM, S-gredzens atrodas gramnegatīvo baktēriju periplazmatiskajā telpā vai grampozitīvo baktēriju peptidoglikāna maisiņā. Divi ārējie gredzeni (D un L) ir neobligāti kustībai, jo tie atrodas tikai gramnegatīvās baktērijās un ir lokalizēti attiecīgi peptidoglikāna slānī un CL ārējā membrānā. Gredzeni S, D un L ir nekustīgi un kalpo, lai fiksētu karogu CS. Kaņģa rotāciju nosaka šūnas CPM iebūvētā M-gredzena rotācija. Tādējādi flagellum bazālā ķermeņa strukturālās iezīmes nosaka CS struktūra.
Funkcionāli bazālais korpuss ir elektromotors, ko darbina protoni. Bāzes korpusa M-gredzenu (rotējošo rotoru) ieskauj membrānas proteīni ar negatīviem lādiņiem (motora stators). Baktēriju šūnai ir efektīvs mehānisms elektroķīmiskās enerģijas pārvēršanai mehāniskajā enerģijā. Tāpēc baktērija tērē apmēram 0,1% no visas enerģijas, ko tā tērē ziepju darbībai. Zogai darbojoties, tiek izmantots protonu dzinējspēks, ko nodrošina protonu koncentrācijas atšķirība membrānas ārējā un iekšējā pusē (ārpusē to ir vairāk) un negatīvāka lādiņa klātbūtne. membrānas iekšējā pusē. Protonu dzinējspēks liek protoniem iziet cauri bazālajam ķermenim šūnā, kamēr tie kavējas noteiktās rotora daļās, dodot tiem pozitīvu lādiņu, pēc tam protoni nonāk šūnā. Uzlādētās sekcijas atrodas tā, ka starp uzlādētajām rotora sekcijām un statoru rodas pievilcīgs spēks, M-gredzens sāk griezties ar ātrumu aptuveni 300 apgr./min. Rotācijas mehānisms: COOH grupas uzlāde-uzlādēšana aminoskābēs. Lai gredzens pilnībā apgrieztos, caur bazālo ķermeni ir jāiziet 500–1000 protonu. M-gredzena griešanās caur asi, kas ir stingri savienota ar to un āķi, tiek pārnesta uz flagellum pavedienu, kas darbojas kā dzenskrūve vai kuģa dzenskrūve. Baktērija peld tik ilgi, kamēr darbojas skrūve, inerces ieguldījums ir ārkārtīgi mazs.
Turklāt baktērijas, pat mirušas, ūdens vidē pārvietojas Brauna kustības rezultātā. Baktēriju šūna pastāvīgi tiek pakļauta apkārtējo molekulu ietekmei, kas atrodas termiskā kustībā. Sitieni no dažādiem virzieniem met baktēriju no vienas puses uz otru.
Frogellas kustības veids ir rotācijas. Ir divu veidu kustības: taisnvirziena un krītoša (periodiskas nejaušas kustības virziena izmaiņas). Kad flagellas griežas pretēji pulksteņrādītāja virzienam (apmēram 1 sekundi) ar frekvenci 40–60 apgr./min (tuvu vidējam elektromotora ātrumam), to pavedieni tiek savīti vienā saišķī (9.a att.). Flagela rotācija tiek pārnesta uz šūnu. Tā kā šūna ir daudz masīvāka par flagellum, tā sāk kustēties taisnā līnijā pretējā virzienā ar ātrumu 3 reizes mazāks par flagellum ātrumu.
Tas nodrošina šūnas translācijas kustību, kuras ātrumu šķidrā vidē dažādi veidi baktērijas ir 20–200 μm/s (tas atbilst aptuveni 300–3000 ķermeņa garumiem minūtē) un lēnāka kustība uz cietās vides virsmas.
Baktērija var mērķtiecīgi peldēt vienā virzienā ne ilgāk kā 3 s, tad apkārtējo molekulu ietekme to pagriež nejaušā virzienā. Tāpēc baktērijas ir izstrādājušas mehānismu spontānai kustības virziena maiņai - flagella motora pārslēgšanai. Kad tā sāk griezties pulksteņrādītāja virzienā (apmēram 0,1 s), baktērija apstājas un apgriežas (veic "sagriešanos") nejaušā virzienā. Šajā gadījumā flagellas izkliedējas dažādos virzienos (9.b att.). Amfitrichā, pārvietojoties, viens flagellas saišķis tiek apgriezts ar iekšpusi uz āru (kā lietussargs, ko vējš apgriež iekšā). Tad motors atkal pārslēdzas uz griešanos pretēji pulksteņrādītāja virzienam, un baktērija atkal peld taisnā līnijā, bet citā, nejaušā virzienā.
Ziedonis var arī mainīt virzienu, reaģējot uz ārēju stimulu. Ja baktērija virzās uz optimālo atraktanta koncentrāciju, flagellas izspiež šūnu caur barotni, tās taisnvirziena kustība kļūst garāka, un buljona frekvence ir zemāka, kas galu galā ļauj tai pārvietoties vēlamajā virzienā.
Ir zināmi neaktīvu (paralizētu) flagella esamības gadījumi. Lai pārvietotos ar baktērijām ar kauliņiem, ir nepieciešama CS integritāte (neskartība). Šūnu apstrāde ar lizocīmu, kas noved pie CS peptidoglikāna slāņa noņemšanas, izraisa baktēriju pārvietošanās spējas zudumu, lai gan flagellas paliek neskartas.
Baktēriju taksometri. Uz redzēšanos Vide paliek nemainīgs, baktērijas peld nejauši. Tomēr vide reti ir pilnīgi viendabīga. Ja vide ir neviendabīga, baktērijām ir elementāras uzvedības reakcijas: tās aktīvi pārvietojas noteiktu ārējo faktoru noteiktajā virzienā. Šādas ģenētiski noteiktas mērķtiecīgas baktēriju kustības sauc par taksometriem. Atkarībā no faktora izšķir ķemotaksi (īpašs gadījums ir aerotakss), fototaksi, magnetotaksi, termotaksi un viskozitaksi.
Ķīmijtakss- kustība noteiktā virzienā attiecībā pret ķīmiskās vielas avotu. Ķīmiskās vielas iedala divās grupās: inertie un izraisošie taksometri – ķīmijefektori. Ķīmijefektoru vidū ir vielas, kas piesaista baktērijas – atraktanti (cukuri, aminoskābes, vitamīni, nukleotīdi), un vielas, kas tās atgrūž – repelenti (dažas aminoskābes, spirti, fenoli, neorganiskie joni). Aerobo atraktants un anaerobo prokariotu repelents ir molekulārais skābeklis. Atraktantus bieži pārstāv pārtikas substrāti, lai gan ne visas organismam nepieciešamās vielas darbojas kā atraktanti. Tāpat ne visas indīgās vielas kalpo kā repelenti un ne visi repelenti ir kaitīgi. Tādējādi baktērijas nespēj reaģēt ne uz kādiem savienojumiem, bet tikai uz noteiktiem un dažādām baktērijām atšķirīgiem.
Baktērijas šūnas virsmas struktūrās atrodas īpašas proteīna molekulas - receptori, kas specifiski savienojas ar noteiktu ķīmijefektoru, savukārt ķīmijefektora molekula nemainās, bet receptora molekulā notiek konformācijas izmaiņas. Receptori atrodas nevienmērīgi pa visu šūnas virsmu un ir koncentrēti vienā no poliem. Receptora stāvoklis atspoguļo attiecīgā efektora ekstracelulāro koncentrāciju.
Chemotaksei ir adaptīva vērtība. Piemēram, holēras vibrio formas ar traucētu ķemotaksiju ir mazāk virulentas.
Aerotaxis- Baktērijas, kurām nepieciešams molekulārais skābeklis, uzkrājas ap gaisa burbuļiem, kas nokrituši zem pārklājuma.
Fototaksis- kustība pret gaismu vai prom no tās, kas raksturīga fototrofiskām baktērijām, kuras izmanto gaismu kā enerģijas avotu.
magnetotaksis- ūdens baktēriju, kas satur dzelzi saturošu minerālu kristālus, spēja peldēt pa Zemes magnētiskā lauka līnijām.
Termotaksē- kustība temperatūras izmaiņu virzienā, kam ir liela nozīme dažām patogēnām baktērijām.
Viskozitaksi- spēja reaģēt uz šķīduma viskozitātes izmaiņām. Baktērijas parasti tiecas uz barotni ar augstāku viskozitāti, kas ir ļoti svarīga patogēnām sugām.
Slīdošās baktērijas. Dažiem prokariotiem, piemēram, mikoplazmām, ir konstatēta spēja slīdēt ar mazu ātrumu (2–11 µm/s) pa cietu vai viskozu substrātu.
Ir vairākas hipotēzes, kas izskaidro slīdošo kustību. Saskaņā ar reaktīvās dzinējspēka hipotēze tas ir saistīts ar gļotu izdalīšanos caur daudzām gļotu porām SC, kā rezultātā šūna tiek atgrūsta no substrāta virzienā, kas ir pretējs gļotu sekrēcijas virzienam. Saskaņā ar ceļojošā viļņa hipotēze slīdošā kustība kustīgās formās bez flagellates ir saistīta ar plānas sakārtotu fibrilu proteīna slāņa klātbūtni starp peptidoglikāna slāni un CL ārējo membrānu, kas ir līdzīga flagellas pavedieniem. Fibrilu rotācijas kustība, ko “palaida” šīs struktūras, noved pie “ceļojoša viļņa” (kustīgu mikroskopisku CL izciļņu) parādīšanās uz šūnas virsmas, kā rezultātā šūna tiek atgrūsta no substrāta. Visbeidzot, dažās slīdošās baktērijās ir aprakstītas struktūras, kas atgādina flagellatu formu bazālos ķermeņus.
Flagela funkcijas:
1. Nodrošiniet saķeri — sākuma stadija infekcijas process.
2. Nodrošināt baktēriju kustīgumu.
3. Nosakiet antigēnu specifiku, tas ir H-antigēns.
Ziedu noteikšana:
1. Dabisko preparātu fāzes kontrasta mikroskopija (“sasmalcināti” un “karinātie” pilieni). Mikroskopiski mobilitāti nosaka ikdienas kultūras šūnās. Lai mobilitāti atšķirtu no pasīvās Brauna kustības, pētāmās kultūras pilienam pievieno pilienu 5% fenola ūdens šķīduma, šajā gadījumā aktīvā kustība apstājas.
2. Dabisko preparātu tumšā lauka mikroskopija.
3. Ar krāsvielām vai metāliem iekrāsotu preparātu gaismas mikroskopija. Tā kā preparāta pagatavošanas laikā flagellas tiek ļoti viegli sabojātas, ikdienas praksē šīs metodes tiek izmantotas reti.
Flagellas iekrāso, izmantojot šūnas, kas audzētas uz agara slīpuma. Šūnas, kas atrodas netālu no kondensācijas ūdens, tiek atlasītas ar baktēriju cilpu un rūpīgi pārnestas uz sterilu destilētu ūdeni, kura temperatūra ir tāda pati kā baktēriju inkubācijas temperatūra uz slīpā agara, un baktērijas netiek izkratas no cilpas, bet gan rūpīgi iegremdētas ūdenī. Caurule ar baktērijām tiek atstāta istabas temperatūrā 30 minūtes. Izmanto ķīmiski tīru (mazgātu hroma maisījumā) stiklu, uz kura uzpilina 2-3 pilienus suspensijas. Suspensija tiek izkliedēta pa stikla virsmu, to viegli noliecot. Izžāvējiet zāles gaisā.
Ziedi ir ļoti plāni, tāpēc tos var noteikt tikai ar īpašu apstrādi. Sākotnēji ar kodināšanas palīdzību tiek panākts pietūkums un to izmēra palielināšanās, un pēc tam preparāts tiek krāsots, kā rezultātā tie kļūst redzami gaismas mikroskopijā.
Biežāk lietots sudrabošanas metode pēc Morozova (10. att.):
- preparātu 1 minūti fiksē ar ledus etiķskābes šķīdumu, mazgā ar ūdeni;
- uz 1 minūti uzklāt tanīna šķīdumu (iedegums, padarot flagellas blīvākus), nomazgāt ar ūdeni;
- preparātu apstrādā, karsējot ar impregnējošu sudraba nitrāta šķīdumu 1-2 minūtes, mazgā ar ūdeni, žāvē un mikroskopē.
Mikroskopiski ir redzamas tumši brūnas šūnas un gaišākas flagellas.

Rīsi. 10. Ziedu identificēšana ar sudrabošanu
Rīsi. vienpadsmit. Ziedu identifikācija
ar elektronu mikroskopu
4. att. Ar smagajiem metāliem nogulsnēto preparātu elektronmikroskopija (11. att.).
5. Netieši - atbilstoši baktēriju augšanas veidam, sējot pusšķidrā 0,3% agarā. Pēc kultūraugu inkubācijas termostatā 1–2 dienas tiek atzīmēts baktēriju augšanas raksturs:
- nekustīgās baktērijās (piem. S.saprophyticus) gar injekciju ir augšana - “nagla”, un barotne ir caurspīdīga;
kustīgās baktērijās (piem. E. coli) ir augšana uz injekcijas pusi, visā agara kolonnā - "siļķe", un barotnes difūzs duļķainums.
Visas baktērijas sadalīts mobilajos un nekustīgajos. Kustības orgāni baktērijās ir flagellas. Tos veido flagellīns, proteīns, kas strukturāli ir saistīts ar miozīna tipa kontraktiliem proteīniem.
Flagellum bāze ir bazāls ķermenis, kas sastāv no disku sistēmas (blefaroplasts: 1 disks - šūnas sienas ārējā puse, 2 disks - šūnas sienas iekšējā puse, 3 diski - citoplazmas membrāna), kas "iegulti" citoplazmā. membrāna un šūnu siena. Kaņģa garums ir lielāks par paša mikroba ķermeņa garumu.
Saskaņā ar flagella skaitu un to atrašanās vietu, kustīgos mikroorganismus iedala:
1. Vienveidīgs, kam ķermeņa galā ir viens flagellum (viskustīgākais). Piemēram, Vibrio cholerae.
2. Lophotrichous, kam pie viena no šūnas poliem ir karogs. Piemēram, Burkholderia (Pseudomonas) pseudomalei ir melioidozes izraisītājs.
3. Amphitrichous, kam abos šūnas polos ir karogs. Piemēram, Spirillum volutans.
4. Peritrichous, kam ir flagellas visā šūnas perimetrā. Piemēram, Escherichia coli, Salmonella typhi.
Flagelu noteikšana. Ziedi ir ļoti plāni, tāpēc tos var noteikt tikai ar īpašu apstrādi. Jo īpaši vispirms ar kodinātāja palīdzību tiek panākts pietūkums un to lieluma palielināšanās, un pēc tam preparāts tiek krāsots, kā rezultātā tie kļūst redzami gaismas mikroskopijā. Flagellas var noteikt, krāsojot pēc Morozova, Leflera, kā arī ar elektronu mikroskopu. Flagella var noteikt arī pēc baktēriju aktīvās kustības.
Mikrobu kustība tiek novērota "sasmalcinātu" un "piekārtu" pilienu preparātos no dzīvām kultūrām. Šie preparāti tiek mikroskopēti ar sausu vai iegremdējamu lēcu tumšā laukā vai fāzes kontrastā. Turklāt kustīgumu var noteikt pēc baktēriju augšanas veida pusšķidrā agarā.
Dzēra no baktērijām.
Pili (pili), sinonīmi: villi, fimbria, ir plāni, dobi proteīna pavedieni, kas pārklāj baktēriju šūnu virsmu. Atšķirībā no flagellas, tās neveic motora funkciju.
Pili stiepjas no šūnas virsmas un sastāv no olbaltumvielām pilina.
Pēc funkcionālā mērķa tie ir sadalīti 2 veidos.
1) Pirmā tipa pilīši atrodas lielākajā daļā baktēriju, tāpēc tos sauc par "vispārējā tipa bārkstiņām" (parasti pili). Tie izraisa baktēriju pieķeršanos vai adhēziju noteiktām saimniekorganisma šūnām. Adhēzija ir jebkura infekcijas procesa sākuma stadija.
2) Otrā tipa pili (sinonīmi: konjugatīvs vai seksuāls - dzimuma pili) ir pieejamas tikai donorbaktērijās, kurām ir īpaša plazmīda. To skaits ir neliels - 1-4 šūnā.
Grīdas zāģi veic šādas funkcijas:
1. Piedalīties ģenētiskā materiāla pārnešanā no vienas šūnas uz otru baktēriju konjugācijas laikā.
2. Tie adsorbē specifiskus baktēriju vīrusus – bakteriofāgus
Baktēriju sporas, veidošanās apstākļi, Aujeski iekrāsošanās vieta, mehānisms un stadijas.
strīds- savdabīga miera baktēriju forma ar grampozitīvu šūnu sienas struktūras veidu.
sporulācija- tas ir sugas (genofora) saglabāšanas veids ārējā vidē nelabvēlīgos apstākļos, nevis pavairošanas metode.
Veidojas sporas baktēriju pastāvēšanai nelabvēlīgos apstākļos (žāvēšana, barības vielu trūkums utt.). Baktēriju šūnas iekšpusē veidojas viena spora (endospora).
Sporulācijas stadijas
1. Sagatavošanas. Baktēriju citoplazmā veidojas sablīvēta zona, kurā nav brīva ūdens, ko sauc par "sporogēno zonu", kurā atrodas nukleoīds.
2. Presporu (prosporu) stadija. Ap sporogēno zonu veidojas dubultās citoplazmas membrānas apvalks.
3. Garozas veidošanās, kas sastāv no peptidoglikāna un ārējās membrānas ar augstu kalcija sāļu un lipīdu saturu.
4. Nogatavināšanas stadija. Ārējās membrānas ārējā pusē veidojas sporas apvalks, pēc kura šūnas veģetatīvā daļa tiek lizēta, atbrīvojot sporu.
Mikrobioloģijas attīstība pēdējās desmitgadēs ir nesusi daudzus atklājumus. Un viena no tām ir kažokādu baktēriju kustības īpatnības. Šo seno organismu dzinēju dizains izrādījās ļoti sarežģīts un saskaņā ar to darbības principu ļoti atšķiras no mūsu tuvāko vienšūņu eikariotu radinieku karogiem. Flagellate baktērijas dzinējspēks ir bijis karstākais strīds starp kreacionistiem un evolucionistiem. Par baktērijām, to karogu motoriem un daudz ko citu - šis raksts.
Vispārējā bioloģija
Sākumā atcerēsimies, kādi organismi tie ir un kādu vietu tie ieņem mūsu planētas organiskās pasaules sistēmā. Baktēriju domēns apvieno milzīgu skaitu vienšūnu prokariotu (bez izveidota kodola) organismu.
Šīs dzīvās šūnas parādījās uz dzīves skatuves gandrīz pirms 4 miljardiem gadu un bija pirmie planētas kolonisti. Tiem var būt ļoti dažādas formas (koki, stieņi, vibrioni, spirohetas), taču lielākā daļa no tām ir ar karogiem.
Kur dzīvo baktērijas? Visur. Uz planētas dzīvo vairāk nekā 5 × 10 30. 1 gramā augsnes ir aptuveni 40 miljoni to, mūsu ķermenī dzīvo līdz 39 triljoniem. Tos var atrast Marianas tranšejas dibenā, karstos "melnos smēķētājus" okeānu dibenā, Antarktīdas ledū, un šobrīd uz jūsu rokām ir līdz pat 10 miljoniem baktēriju.
Vērtība ir nenoliedzama
Neskatoties uz to mikroskopisko izmēru (0,5–5 mikroni), to kopējā biomasa uz Zemes ir lielāka nekā dzīvnieku un augu biomasa kopā. To loma vielu apritē ir neaizstājama, un to patērētāju (organisko vielu iznīcinātāju) īpašības neļauj planētu pārklāt ar līķu kalniem.
Nu, neaizmirstiet par patogēniem: mēra, baku, sifilisa, tuberkulozes un daudzu citu infekcijas slimību izraisītāji ir arī baktērijas.
Baktērijas ir atradušas pielietojumu cilvēka saimnieciskajā darbībā. Sākot ar pārtikas rūpniecību ( piena produkti, sieri, marinēti dārzeņi, alkoholiskie dzērieni), "zaļo" ekonomiku (biodegvielu un biogāzi) uz šūnu inženierijas metodēm un zāļu (vakcīnas, serumi, hormoni, vitamīni) ražošanu.

Vispārējā morfoloģija
Kā jau minēts, šiem vienšūnu dzīvības pārstāvjiem nav kodola, to iedzimtais materiāls (DNS molekulas gredzena formā) atrodas noteiktā citoplazmas (nukleoīda) apgabalā. Viņu šūnai ir plazmas membrāna un blīva kapsula, ko veido peptidoglikāna mureīns. No šūnu organellām baktērijām ir mitohondriji, var būt hloroplasti un citas struktūras ar dažādām funkcijām.
Lielākā daļa baktēriju ir flagellas. Cieši esošā kapsula uz šūnas virsmas neļauj tām pārvietoties, mainot pašu šūnu, kā to dara amēba. Viņu flagellas ir dažāda garuma un apmēram 20 nm diametrā blīvi proteīnu veidojumi. Dažām baktērijām ir viens flagellum (monotrichous), bet citām ir divi (amfitrichous). Dažreiz flagellas ir sakārtotas saišķos (lophotrichous) vai aptver visu šūnas virsmu (peritrichous).
Daudzi no tiem dzīvo kā vienas šūnas, bet daži veido kopas (pārus, ķēdes, pavedienus, hifus).

Kustību īpašības
Karogotās baktērijas var pārvietoties dažādos veidos. Daži virzās tikai uz priekšu un maina virzienu, krītot. Daži spēj raustīties, citi pārvietojas, slīdot.
Baktēriju flagellas pilda ne tikai šūnu "aiļa" funkcijas, bet var būt arī "iekāpšanas" rīks.
Vēl nesen tika uzskatīts, ka baktērijas flagellum vicinās kā čūskai aste. Nesenie pētījumi liecina, ka baktēriju kauliņš ir daudz sarežģītāks. Tas darbojas kā turbīna. Piestiprināts pie piedziņas, tas griežas vienā virzienā. Baktēriju izpildmehānisms jeb karogdziedzera motors ir sarežģīta molekulāra struktūra, kas darbojas kā muskuļi. Ar atšķirību, ka muskulim pēc kontrakcijas ir jāatslābinās, un baktēriju motors strādā pastāvīgi.

Kaņģa nanomehānisms
Neiedziļinoties kustību bioķīmijā, atzīmējam, ka flagellum drive izveidē ir iesaistīti līdz 240 proteīniem, kas sadalīti 50 molekulāros komponentos ar noteiktu funkciju sistēmā.
Šajā baktēriju piedziņas sistēmā ir rotors, kas kustas, un stators, kas nodrošina šo kustību. Ir piedziņas vārpsta, bukse, sajūgs, bremzes un akseleratori
Šis miniatūrais dzinējs ļauj baktērijām pārvietoties 35 reizes vairāk nekā tās pašas tikai 1 sekundē. Tajā pašā laikā paša karogs, kas veic 60 tūkstošus apgriezienu minūtē, ķermenis tērē tikai 0,1% no visas šūnas patērētās enerģijas.
Pārsteidzoši ir arī tas, ka baktērija var nomainīt un salabot visas sava motora mehānisma rezerves daļas “ceļā”. Iedomājieties, ka atrodaties lidmašīnā. Un tehniķi maina darbojoša motora asmeņus.
Flagellate baktērija pret Darvinu
Dzinējs, kas spēj darboties ar ātrumu līdz 60 000 apgriezieniem minūtē, pats iedarbinās un kā degvielu izmanto tikai ogļhidrātus (cukuru), kuram ir elektromotoram līdzīga ierīce - vai tāda ierīce varēja rasties evolūcijas procesā?
Šo jautājumu 1988. gadā sev uzdeva bioloģijas zinātņu doktors Maikls Behe. Viņš bioloģijā ieviesa nesamazināmas sistēmas jēdzienu - sistēmu, kurā visas tās daļas ir vienlaikus nepieciešamas, lai nodrošinātu tās darbību, un vismaz vienas daļas noņemšana noved pie tās darbības pilnīgas pārtraukšanas.
No Darvina evolūcijas viedokļa visas strukturālās izmaiņas organismā notiek pakāpeniski, un tikai veiksmīgās tiek atlasītas dabiskās atlases ceļā.
M. Behes secinājumi, kas izklāstīti grāmatā "Darvina melnā kaste" (1996): karogbaktērijas dzinējs ir nedalāma sistēma, kas sastāv no vairāk nekā 40 daļām, un vismaz vienas daļas trūkums novedīs pie pilnīgas disfunkcijas. sistēma, kas nozīmē, ka šī sistēma nevarēja rasties dabiskās atlases ceļā.

Balzams kreacionistiem
Radīšanas teorija, ko iepazīstināja zinātnieks un bioloģijas profesors, Betlēmes Lehigas universitātes (ASV) Bioloģijas zinātņu fakultātes dekāns M. Behe, nekavējoties piesaistīja baznīcas kalpotāju un dievišķā teorijas atbalstītāju uzmanību. dzīvības izcelsme.
2005. gadā Amerikas Savienotajās Valstīs pat notika tiesas prāva, kurā Behe bija liecinieks no "saprātīga dizaina" teorijas piekritējiem, kurā tika apsvērta kreacionisma studiju ieviešana Doveras skolās kursā " Par pandām un cilvēkiem." Process tika zaudēts, šāda priekšmeta mācīšana tika atzīta par pretēju spēkā esošajai konstitūcijai.
Bet debates starp kreacionistiem un evolucionistiem turpinās arī šodien.
Karogs ir baktēriju šūnas virsmas struktūra, kas kalpo tām kustībai šķidrā vidē.
Atkarībā no flagellas atrašanās vietas baktērijas iedala (1. att.):
Peritrihiāls
sajaukts
Pole
Subpolārs
Pole flagella- viena vai vairākas flagellas atrodas uz viena (monopolāra) vai abiem (bipolāriem) šūnas poliem un pamatne ir paralēla šūnas garajai asij.
Subpolāri flagellas(subpolāri) - viens vai vairāki flagellas atrodas sānu virsmas pārejas punktā uz šūnas polu vienā vai divos tās galos. Pamatnē ir taisns leņķis ar šūnas garo asi.
Sānu flagellas(sānu) - vienas vai vairākas kauliņi kūlīša veidā atrodas vienas šūnas pusītes viduspunktā.
Peritrichial flagellas- atrodas pa visu šūnas virsmu pa vienam vai saišķos, stabiem parasti tie tiek atņemti.
Jauktas flagellas- atrodas divas vai vairākas flagellas dažādi punktišūnas.
Atkarībā no flagellu skaita izšķir:
Monotrichous - viens flagellum
Politrihs - ķekars flagellas
Atšķiras arī:
lophotrichous- flagellas monopolitisks politrihiāls izvietojums.
amfitrihi- flagellas bipolārais politrihiāls izvietojums.
Baktēriju flagellum un bazālā ķermeņa struktūra. Flagellum.
Pati flagellum ir sakārtots pavisam vienkārši: kvēldiegs, kas piestiprināts pie pamatķermeņa. Dažreiz starp pamatkorpusu un kvēldiegu var ievietot izliektu caurules daļu, tā saukto āķi; tas ir biezāks par kvēldiegu un piedalās kvēldiega elastīgā piestiprināšanā pie pamatnes.
Ķīmiskā sastāva ziņā flagellum sastāv no 98% flagellīna proteīna (flagellum - flagellum), tajā ir 16 aminoskābes, dominē glutamīns un asparagīnskābes, nelielā daudzumā aromātisko aminoskābju nav triptofāna, cisteīna un cistīna. Flagellīns uzliek antigēnu specifiskumu, to sauc par H-antigēnu. Baktēriju flagellām nav ATPāzes aktivitātes.
Kaņģa biezums ir 10–12 nm, garums 3–15 µm.
Tā ir stingra spirāle, kas savīta pretēji pulksteņrādītāja virzienam. Karogs tiek griezts arī pretēji pulksteņrādītāja virzienam ar frekvenci no 40 apgr./min līdz 60 apgr./min, kas liek šūnai griezties pretējā virzienā, bet kopš tā laika šūna ir daudz smagāka par flagellum, tad tās rotācija ir lēnāka no 12 līdz 14 apgr./min.
Karogs aug no distālā gala, kur apakšvienības iekļūst caur iekšējo kanālu. Dažām sugām flagellum no ārpuses papildus ir pārklāts ar apvalku, kas ir šūnas sienas turpinājums un, iespējams, ir tāda pati struktūra.
Bāzes ķermenis
Bāzes korpuss sastāv no 4 daļām:
Stieņu pārošanās ar kvēldiegu vai āķi
Uz stieņa savērti divi diski. (M un S)
Olbaltumvielu kompleksu grupa (statori)
proteīna vāciņš
Baktērijām, kurām ir iekšējā un ārējā membrāna, ir 2 papildu diski (P un L) un proteīnu struktūras, kas atrodas uz ārējās membrānas pie bazālā ķermeņa, tāpēc tām nav svarīgas kustības.
Bāzes ķermeņa uzbūves īpatnību nosaka šūnas sienas uzbūve: tās neskartums ir nepieciešams flagellas kustībai. Šūnu apstrāde ar lizocīmu noved pie peptidoglikāna slāņa noņemšanas no šūnu sienas, kas noved pie kustību zuduma, lai gan netika traucēta kauliņa struktūra.
Baktēriju flagellas nosaka baktēriju šūnas kustīgumu. Flagella ir plāni pavedieni, kas nāk no citoplazmas membrānas un ir garāki par pašu šūnu. Ziedi ir 12–20 nm biezi un 3–15 µm gari. Tie sastāv no 3 daļām: spirālveida vītnes, āķa un pamata korpusa, kas satur stieni ar īpašiem diskiem (1 disku pāris grampozitīvām un 2 disku pāri gramnegatīvām baktērijām). Ziedu diski ir piestiprināti pie citoplazmas membrānas un šūnas sienas. Tas rada elektromotora efektu ar motora stieni, kas rotē flagellum. Flagella sastāv no proteīna - flagellīna (no flagellum - flagellum); ir H antigēns. Flagellina apakšvienības ir satītas. Ziedu skaits dažādu sugu baktērijās svārstās no viena (monotrich) Vibrio cholerae līdz desmit vai simtiem flagellas, kas stiepjas gar baktērijas (peritrich) perimetru Escherichia coli, Proteus utt. šūnas beigas. Amphitrichous šūnas pretējos galos ir viens flagellum vai zibens saišķis.
Pili (fimbriae, villi) - pavedienveida veidojumi, tievāki un īsāki (3-10 nm x 0,3-10 mikroni) nekā flagellas. Pili stiepjas no šūnas virsmas un sastāv no pilīna proteīna, kam ir antigēna aktivitāte. Ir pili, kas atbild par adhēziju, tas ir, par baktēriju pievienošanu skartajai šūnai, kā arī pili, kas ir atbildīgi par uzturu, ūdens-sāls metabolismu un seksuālo (F-pili), vai konjugācijas pili. Dzērieni ir daudz - vairāki simti katrā būrī. Tomēr dzimuma pili parasti ir 1-3 vienā šūnā: tos veido tā sauktās "vīriešu" donoru šūnas, kas satur transmisīvās plazmīdas (F-, R-, Col-plazmīdas). Dzimuma pili atšķirīga iezīme ir mijiedarbība ar īpašiem "vīriešu" sfēriskiem bakteriofāgiem, kas intensīvi adsorbējas uz dzimuma pili.
Sporas ir savdabīga snaudošu firmicute baktēriju forma, t.i. baktērijas ar grampozitīvu šūnu sienas struktūru. Sporas veidojas baktēriju eksistencei nelabvēlīgos apstākļos (žāvēšana, barības vielu deficīts utt. Baktērijas šūnas iekšienē veidojas viena spora (endospora) Sporu veidošanās veicina sugas saglabāšanos un nav vairošanās metode , tāpat kā sēnēs.Bacillus ģints sporas veidojošajām baktērijām ir sporas, kas nepārsniedz šūnas diametru.Baktērijas, kuru sporu izmērs pārsniedz šūnas diametru, sauc par klostrīdijām, piemēram, par Clostridium ģints baktērijām (lat. Clostridium - vārpstiņa).Sporas ir skābes izturīgas, tāpēc tās tiek iekrāsotas sarkanā krāsā pēc Aujeski metodes vai pēc Ziehl-Nelsen metodes, bet veģetatīvā šūna - zilā krāsā.
Strīda forma var būt ovāla, sfēriska; atrašanās vieta šūnā ir termināla, t.i. nūjas galā (stingumkrampju izraisītājā), apakšgalā - tuvāk nūjas galam (botulisma, gāzes gangrēnas patogēnos) un centrālajā (sibīrijas mēra baciļos). Sporas saglabājas ilgu laiku, jo ir daudzslāņu apvalks, kalcija dipikolināts, zems ūdens saturs un lēni vielmaiņas procesi. Labvēlīgos apstākļos sporas dīgst trīs secīgos posmos: aktivācija, iniciācija, dīgšana.
8. Galvenās baktēriju formas
Globulāras baktērijas (koki) parasti ir sfēriskas, bet var būt nedaudz ovālas vai pupiņas formas. Cocci var atrasties atsevišķi (mikrokoki); pa pāriem (diplokoki); ķēžu (streptokoku) vai vīnogu ķekaru (stafilokoku) veidā, iepakojumā (sarcinas). Streptokoki var izraisīt tonsilītu un erysipelas, stafilokoki – dažādus iekaisuma un strutojošus procesus.
stieņa formas baktērijas visbiežāk. Stieņi var būt atsevišķi, savienoti pa pāriem (diplobaktērijas) vai ķēdēs (streptobaktērijas). Pie nūjiņveidīgajām baktērijām pieder Escherichia coli, salmonelozes, dizentērijas, vēdertīfa, tuberkulozes u.c. patogēni. Dažām stieņveida baktērijām ir iespēja veidoties nelabvēlīgos apstākļos. strīdi. Sporas veidojošos stieņus sauc baciļi. Tiek saukti vārpstveida baciļi klostridijas.
Sporulācija ir sarežģīts process. Sporas būtiski atšķiras no parastās baktēriju šūnas. Viņiem ir blīvs apvalks un ļoti mazs ūdens daudzums, tiem nav vajadzīgas barības vielas, un vairošanās pilnībā apstājas. Sporas ilgstoši spēj izturēt izžūšanu, augstu un zemu temperatūru un dzīvotspējīgā stāvoklī var atrasties desmitiem un simtiem gadu (sibīrijas mēra, botulisma, stingumkrampju u.c. sporas). Nonākušas labvēlīgā vidē, sporas izdīgst, tas ir, pārvēršas ierastajā veģetatīvās pavairošanas formā.
Sarežģītas baktērijas var būt komata formā - vibrios, ar vairākām cirtām - spirilla, plānas savītas nūjas formā - spirohetas. Holēras izraisītājs ir vibrācijas, un sifilisa izraisītājs ir spiroheta.
9. Riketsijas un hlamīdijas morfoloģijas pazīmes
Riketijas ir mazi gramnegatīvi mikroorganismi, kam raksturīgs izteikts polimorfisms – tie veido kokus, stieņveida un pavedienveida formas (22. att.). Riketsiju izmēri svārstās no 0,5 līdz 3-4 mikroniem, pavedienu formu garums sasniedz 10-40 mikronus. Tie neveido sporas un kapsulas, tās pēc Zdrodovska domām ir iekrāsotas sarkanā krāsā.
Hlamīdijas ir sfēriskas, olveida vai stieņa formas. To izmēri svārstās 0,2-1,5 mikronu robežās. Hlamīdiju morfoloģija un lielums ir atkarīgi no to intracelulārās attīstības cikla stadijas, kam raksturīga neliela sfēriska elementāra veidojuma pārvēršanās lielā sākotnējā ķermenī ar bināru dalījumu. Pirms dalīšanas hlamīdijas daļiņas tiek aptvertas veidojumā, kas atgādina baktēriju kapsulu. Hlamīdiju traips "* pēc Romanovska-Giemsa, gramnegatīvs, skaidri redzams intravitālajos preparātos ar fāzes kontrasta mikroskopiju.
10. Mikoplazmu uzbūve un bioloģija.
Tikai viena kārta, Mycoplasmatales, pieder Mollicutes klasei. Šīs kārtas pārstāvji - mikoplazmas -
Tās atšķiras no baktērijām ar to, ka tām trūkst šūnu sienas. Tā vietā tie satur trīs slāņu lipoproteīnu citoplazmas membrānu. Mikoplazmas izmēri svārstās 125-250 mikronu robežās. Tās ir apaļas, ovālas vai pavedienveida, gramnegatīvas.
Mikoplazmas vairojas binārās dalīšanās ceļā, tāpat kā lielākā daļa baktēriju, īpaši pēc nelielu kokosveida veidojumu (elementārķermeņu, EB) veidošanās pavedienveida struktūrās.
Mikoplazmas spēj veidoties un segmentēties. Minimālā reproducēšanas vienība ir ET (0,7-0,2 mikroni). Šūnu membrānas galvenā sastāvdaļa ir holesterīns. Mikoplazmas nespēj veidot holesterīnu un izmantot to no audiem vai uzturvielu barotnēm, kas papildinātas ar to ievadīšanu. Gram-krāsošana ir negatīva, bet Romanovska-Giemsa krāsošana dod vislabākos rezultātus. Mikoplazmas ir prasīgas pret audzēšanas apstākļiem: barības barotnei jāpievieno dabīgais serums, holesterīns, nukleīnskābes, ogļhidrāti, vitamīni un dažādi sāļi. Uz blīvām barotnēm tie veido raksturīgas nelielas caurspīdīgas kolonijas ar izceltu granulu centru, piešķirot tām "ceptu olu" izskatu. Uz barotnēm ar asinīm daži mikoplazmu veidi veic a- un beta-hemolīzi. Pusšķidrā vidē mikoplazmas aug gar injekcijas līniju, veidojot izkliedētas, drupanas kolonijas. Šķidrā vidē tie rada nelielu miglainību vai opalescenci; daži celmi spēj veidot visplānāko taukaino plēvi. Cilvēkiem ir izolēti Mycoplasma, Ureaplasma un Acholeplasma ģints pārstāvji, tostarp patogēnās un saprofītiskās sugas.