According to the location of the flagella, bacteria are divided into. Organelles of movement: functions and structure, features of the movement of protozoa
Structure. About half of the known species of bacteria on the surface have organs of movement - wave-like curved flagella. The mass of flagella is up to 2% of the dry mass of the bacterium. The length of the flagellum is longer than the length of the body of the microorganism and is 3–12 μm; the thickness of the flagellum is 0.02 μm, and the polar flagella are thicker than the peritrichous ones.
Flagella are composed of flagellin protein (Latin flagella - flagellum), which in its structure belongs to contractile proteins of the myosin type. The flagellum contains either one homogeneous protein filament or 2–3 filaments tightly folded into a braid. The thread of the flagellum is a rigid spiral twisted counterclockwise; the pitch of the helix is specific to each bacterial species.
The number, size, and arrangement of flagella are features that are constant for a particular species and are taken into account in taxonomy. However, some bacteria can form flagella. different types. In addition, the presence of flagella depends on environmental conditions: on solid media during long-term cultivation, bacteria can lose flagella, and on liquid media they can be acquired again. The number and location of flagella in the same species can be determined by the stage of the life cycle. Therefore, the taxonomic significance of this feature should not be overestimated.
Classification of bacteria according to the number and location of flagella:
1. Atrichi - flagella are absent.
2. Monotrichus- one flagellum located at one of the poles of the cell (genus Vibrio)- monopolar monotrichous arrangement of flagella, the most mobile bacteria.
3. Polytrichus - many flagella:
– lophotrichous- bundle of flagella at one pole of the cell (birth Pseudomonas, Burkholderia) - monopolar polytrichous arrangement of flagella;
– amphitrichous- at each pole of the cell there is a bundle of flagellum (genus Spirillum)- bipolar polytrichous arrangement of flagella;
– peritrichous- flagella are arranged in no particular order over the entire surface of the cell (fam. Enterobacteriaceae(birth Escherichia, Proteus), fam. bacillaceae, family Clostidiaceae), the number of flagella varies from 6 to 1000 per cell, depending on the type of bacteria (Fig. 7).
Fig.7. Options for the location of flagella in bacteria:
1 - monotrich, 2 - lophotrich;
3 - amphitrichous; 4 - peritrich.
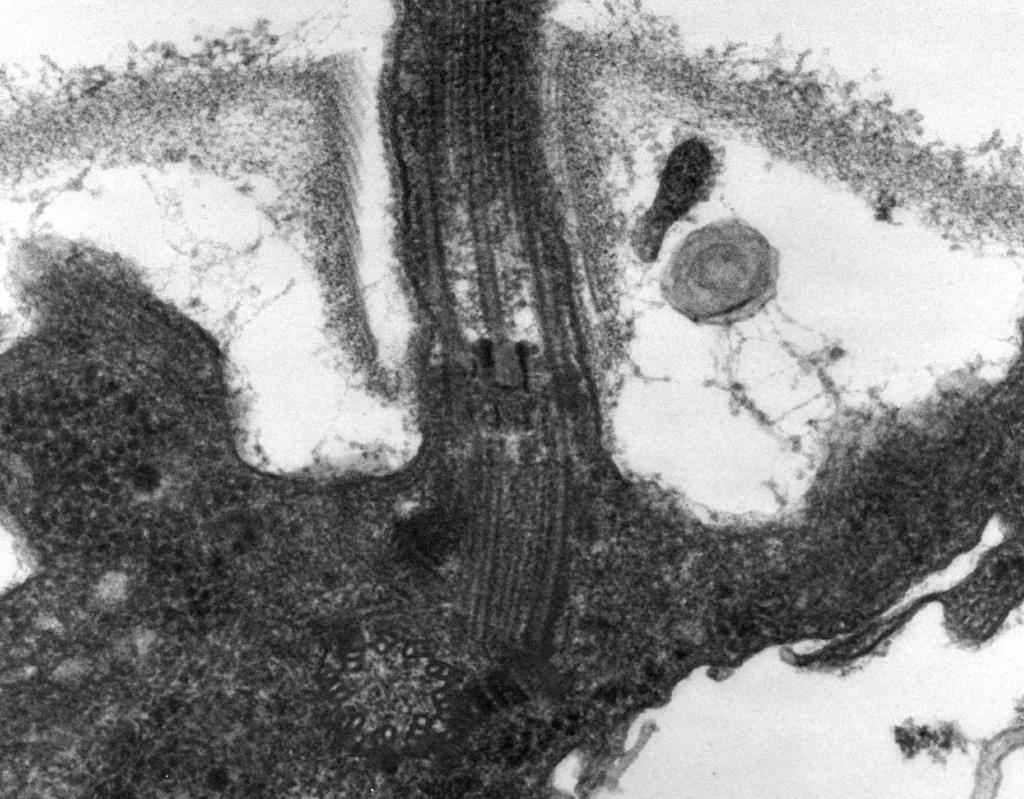
– Electron microscopy reveals that flagellum consists of three parts: spiral filament, hook and basal body (Fig. 8).
The main part of the flagellum is long spiral thread (fibril) - a rigid hollow cylinder with a diameter of about 120 nm, consisting of flagellin protein. Protein molecules form 11 rows along the length of the thread and are arranged in a spiral. During the growth of the thread, protein molecules synthesized inside the cell pass through the cavity of the cylinder and are attached to a spiral at its end. At the end of the flagellum there is a protein cap (lid) that closes the opening of the cylinder and prevents the release of protein molecules into the environment. The length of the flagellum filament can reach several micrometers. In some species of bacteria, the flagellum is additionally covered with a sheath on the outside. At the surface of the CS, the spiral thread passes into a thickened curved structure - a hook.

Rice. eight. Scheme of the structure of the flagellum
2. Hook(20–45 nm thick) near the cell surface is a relatively short cylinder, composed of a protein different from flagellin, and serves to provide a flexible connection of the filament to the basal body.
3. Basal body located at the base of the flagellum and ensures its rotation. The basal body contains 9–12 different proteins and consists of two or four disks (rings) strung on a rod, which is a continuation of the hook. These rings are mounted in the CPM and CS. The two inner rings (M and S) are obligatory components of the basal body. The M-ring is localized in the CPM, the S-ring is located in the periplasmic space of Gram-negative bacteria or in the peptidoglycan sac of Gram-positive bacteria. The two outer rings (D and L) are optional for movement, since they are present only in Gram-negative bacteria and are localized, respectively, in the peptidoglycan layer and in the outer membrane of the CL. Rings S, D, and L are immobile and serve to fix the flagellum in the CS. The rotation of the flagellum is determined by the rotation of the M-ring built into the CPM of the cell. Thus, structural features of the basal body of the flagellum are determined by the structure of the CS.
Functionally, the basal body is an electric motor powered by protons. The M-ring of the basal body (rotating rotor) is surrounded by membrane proteins with negative charges (motor stator). The bacterial cell has an efficient mechanism for converting electrochemical energy into mechanical energy. Therefore, the bacterium spends about 0.1% of all the energy it expends on the operation of the flagellum. When the flagellum operates, a proton-motive force is used, which is provided by the difference in proton concentrations on the outer and inner sides of the membrane (there are more of them on the outer side) and the presence of a more negative charge on the inner side of the membrane. The proton motive force forces the protons to pass through the basal body into the cell, while they linger in certain parts of the rotor, giving them a positive charge, then the protons go inside the cell. The charged sections are located in such a way that an attractive force arises between the charged sections of the rotor and the stator, the M-ring begins to rotate at a speed of about 300 rpm. Rotation mechanism: charge-recharge of the COOH group in amino acids. For a complete revolution of the ring, 500–1000 protons must pass through the basal body. The rotation of the M-ring through the axis rigidly connected to it and the hook is transmitted to the flagellum filament, which functions as a propeller or a ship's propeller. The bacterium swims as long as the screw is working, the contribution of inertia is extremely small.
In addition, bacteria, even dead ones, in the aquatic environment move as a result of Brownian motion. The bacterial cell is constantly subjected to impacts of the surrounding molecules, which are in thermal motion. Hits from different directions throw the bacterium from side to side.
The type of movement of the flagella is rotational. There are two types of movement: rectilinear and tumbling (periodic random changes in direction of movement). When the flagella rotate counterclockwise (about 1 second), at a frequency of 40–60 rpm (close to the speed of an average electric motor), their threads are woven into a single bundle (Fig. 9a). The rotation of the flagella is transmitted to the cell. Since the cell is much more massive than the flagellum, it begins to move in a straight line in the opposite direction, at a speed 3 times less than the speed of the flagellum.
This ensures the translational movement of the cell, the speed of which in a liquid medium for different types bacteria is 20–200 μm/s (this corresponds to about 300–3000 body lengths per minute) and slower movement on the surface of solid media.
The bacterium can swim purposefully in one direction for no more than 3 s, then the impacts of the surrounding molecules turn it in a random direction. Therefore, bacteria have developed a mechanism for a spontaneous change in the direction of movement - switching the flagellar motor. When it begins to rotate clockwise (about 0.1 s), the bacterium stops and flips (performs a “tumble”) in a random direction. In this case, the flagella scatter in different directions (Fig. 9b). In amphitrichous, when moving, one bundle of flagella is turned inside out (like an umbrella turned inside out by the wind). Then the motor again switches to counterclockwise rotation, and the bacterium again swims in a straight line, but in a different, random direction.
The flagella can also change direction in response to an external stimulus. If the bacterium moves towards the optimal attractant concentration, the flagella push the cell through the medium, its rectilinear movement becomes longer, and the tumbling frequency is lower, which ultimately allows it to move in the desired direction.
Cases of the existence of inactive (paralyzed) flagella are known. For the movement of flagellated bacteria, the integrity (intactness) of the CS is necessary. The treatment of cells with lysozyme, which leads to the removal of the peptidoglycan layer of the CS, causes the loss of the bacteria's ability to move, although the flagella remain intact.
Taxis of bacteria. Bye environment remains unchanged, the bacteria swim randomly. However, the environment is rarely completely homogeneous. If the environment is heterogeneous, bacteria exhibit elementary behavioral reactions: they actively move in the direction determined by certain external factors. Such genetically determined purposeful movements of bacteria are called taxis. Depending on the factor, chemotaxis (a special case is aerotaxis), phototaxis, magnetotaxis, thermotaxis and viscositaxis are distinguished.
Chemotaxis- movement in a certain direction relative to the source of the chemical. Chemical substances divided into two groups: inert and causing taxis - chemoeffectors. Among chemoeffectors there are substances that attract bacteria - attractants (sugars, amino acids, vitamins, nucleotides), and substances that repel them - repellents (some amino acids, alcohols, phenols, inorganic ions). The attractant for aerobic and repellent for anaerobic prokaryotes is molecular oxygen. Attractants are often represented by food substrates, although not all substances necessary for the body act as attractants. Also, not all poisonous substances serve as repellents and not all repellents are harmful. Thus, bacteria are not able to react to any compounds, but only to certain and different ones for different bacteria.
In the surface structures of a bacterial cell, there are special protein molecules - receptors that specifically connect to a certain chemoeffector, while the chemoeffector molecule does not change, but conformational changes occur in the receptor molecule. Receptors are located unevenly over the entire surface of the cell, and are concentrated at one of the poles. The state of the receptor reflects the extracellular concentration of the corresponding effector.
Chemotaxis has an adaptive value. For example, forms of cholera vibrio with impaired chemotaxis are less virulent.
Aerotaxis- Bacteria that need molecular oxygen accumulate around air bubbles that have fallen under a coverslip.
Phototaxis- movement towards or away from light, characteristic of phototrophic bacteria that use light as an energy source.
magnetotaxis- the ability of aquatic bacteria containing crystals of iron-containing minerals to swim along the lines of the Earth's magnetic field.
Thermotaxis- movement in the direction of temperature change, which has great importance for some pathogenic bacteria.
Viscositaxis- the ability to respond to changes in the viscosity of the solution. Bacteria usually tend to a medium with a higher viscosity, which is of great importance for pathogenic species.
Sliding bacteria. The ability to slide at a low speed (2–11 µm/s) over a solid or viscous substrate has been found in some prokaryotes, for example, mycoplasmas.
There are several hypotheses that explain the sliding movement. According to jet propulsion hypothesis it is due to the secretion of mucus through numerous mucus pores in the SC, as a result of which the cell is repelled from the substrate in the direction opposite to the direction of mucus secretion. According to the traveling wave hypothesis gliding motion in motile flagellate-free forms is associated with the presence between the peptidoglycan layer and the outer membrane of the CL of a thin protein layer of ordered fibrils, similar to the filaments of flagella. The rotational movement of fibrils, “launched” by these structures, leads to the appearance of a “travelling wave” (moving microscopic bulges of the CL) on the cell surface, as a result of which the cell is repelled from the substrate. Finally, structures resembling the basal bodies of flagellated forms have been described in some gliding bacteria.
Functions of flagella:
1. Provide adhesion - initial stage infectious process.
2. Provide motility of bacteria.
3. Determine the antigenic specificity, this is the H-antigen.
Detection of flagella:
1. Phase contrast microscopy of native preparations (“crushed” and “hanging” drops). Microscopically, the mobility is determined in the cells of the daily culture. In order to distinguish mobility from passive Brownian motion, a drop of a 5% aqueous solution of phenol is added to a drop of the studied culture; in this case, active movement stops.
2. Dark-field microscopy of native preparations.
3. Light microscopy of preparations stained with dyes or metals. Since the flagella are very easily damaged during the preparation of the preparation, these methods are rarely used in everyday practice.
Flagella are stained using cells grown on agar slant. Cells located near the condensation water are selected with a bacterial loop and carefully transferred to sterile distilled water of the same temperature as the temperature of incubation of bacteria on slant agar, and the bacteria are not shaken off the loop, but carefully immersed in water. The tube with bacteria is left at room temperature for 30 minutes. Chemically pure (washed in a chromium mixture) glass is used, on which 2-3 drops of suspension are applied. The suspension is spread over the glass surface by gently tilting it. Air dry the drug.
The flagella are very thin, so they can only be detected with special processing. Initially, with the help of etching, swelling and an increase in their size are achieved, and then the preparation is colored, due to which they become visible under light microscopy.
More often used silvering method according to Morozov (Fig. 10):
- the preparation is fixed with a solution of glacial acetic acid for 1 minute, washed with water;
- apply a solution of tannin (tanning, making the flagella more dense) for 1 minute, washed with water;
- the preparation is treated by heating with an impregnating solution of silver nitrate for 1-2 minutes, washed with water, dried and microscoped.
Microscopically, dark brown cells and lighter flagella are visible.

Rice. ten. Identification of flagella by silvering
Rice. eleven. Identification of flagella
by electron microscopy
Fig. 4. Electron microscopy of preparations deposited with heavy metals (Fig. 11).
5. Indirectly - according to the nature of bacterial growth when sown in semi-liquid 0.3% agar. After incubation of crops in a thermostat for 1–2 days, the nature of bacterial growth is noted:
- in non-motile bacteria (eg. S.saprophyticus) there is growth along the injection - a “nail”, and the medium is transparent;
in motile bacteria (eg. E. coli) there is growth to the side of the injection, throughout the agar column - "herringbone", and diffuse turbidity of the medium.
All bacteria divided into mobile and immobile. The organs of movement in bacteria are flagella. They are made up of flagellin, a protein that is structurally related to myosin-type contractile proteins.
Flagellum base is a basal body, consisting of a system of disks (blepharoplast: 1 disk - the outer side of the cell wall, 2 disk - the inner side of the cell wall, 3 disk - the cytoplasmic membrane), "embedded" in the cytoplasmic membrane and cell wall. The length of the flagellum is greater than the length of the body of the microbe itself.
According to the number of flagella and their location, motile microorganisms are divided into:
1. Monotrichous, having one flagellum at the end of the body (the most mobile). For example, Vibrio cholerae.
2. Lophotrichous, having a bundle of flagella at one of the poles of the cell. For example, Burkholderia (Pseudomonas) pseudomalei is the causative agent of melioidosis.
3. Amphitrichous, having a flagellum at both poles of the cell. For example, Spirillum volutans.
4. Peritrichous, having flagella around the entire perimeter of the cell. For example, Escherichia coli, Salmonella typhi.
Detection of flagella. The flagella are very thin, so they can only be detected with special processing. In particular, at first, with the help of a mordant, swelling and an increase in their size are achieved, and then the preparation is colored, due to which they become visible under light microscopy. Flagella can be detected by staining according to Morozov, Leffler, as well as by electron microscopy. Flagella can also be detected by the active motility of bacteria.
The movement of microbes is observed in preparations of "crushed" and "hanging" drops from living cultures. These preparations are microscoped with a dry or immersion lens in a dark field or in phase contrast. In addition, motility can be determined by the nature of the growth of bacteria in semi-liquid agar.
Drank from bacteria.
Pili (pili), synonyms: villi, fimbria, are thin hollow filaments of protein nature that cover the surface of bacterial cells. Unlike flagella, they do not perform a motor function.
Pili extend from the cell surface and are composed of protein pilina.
According to their functional purpose, they are divided into 2 types.
1) Pili of the first type are present in most bacteria, so they are called "general type villi" (common pili). They cause the attachment or adhesion of bacteria to certain cells of the host organism. Adhesion is the initial stage of any infectious process.
2) Pili of the second type (synonyms: conjugative, or sexual - sex pili) are available only in donor bacteria that have a special plasmid. Their number is small - 1-4 per cell.
Floor saws perform the following functions:
1. Participate in the transfer of genetic material from one cell to another during the conjugation of bacteria.
2. They adsorb specific bacterial viruses - bacteriophages
Bacterial spores, formation conditions, location, mechanism and stages of Aujeszky staining.
controversy- a peculiar form of resting bacteria with a gram-positive type of cell wall structure.
sporulation- this is a way of preserving a species (genophore) in the external environment under adverse conditions, and not a method of reproduction.
Spores are formed under unfavorable conditions for the existence of bacteria (drying, nutrient deficiency, etc.). Inside the bacterial cell, one spore (endospore) is formed.
Stages of sporulation
1. Preparatory. In the cytoplasm of bacteria, a compacted area is formed that does not have free water, called the "sporogenous zone", which contains the nucleoid.
2. Stage of prespores (prospores). Around the sporogenous zone, a sheath of a double cytoplasmic membrane is formed.
3. Formation of a cortex consisting of peptidoglycan and an outer membrane with a high content of calcium salts and lipids.
4. Stage of maturation. A spore envelope is formed on the outer side of the outer membrane, after which the vegetative part of the cell is lysed, releasing the spore.
The development of microbiology has brought many discoveries in recent decades. And one of them is the peculiarities of the movement of flagellated bacteria. The design of the engines of these ancient organisms turned out to be very complex and, according to the principle of their work, is very different from the flagella of our closest eukaryotic relatives of the protozoa. The engine of the flagellate bacterium has been the hottest controversy between creationists and evolutionists. About bacteria, their flagellar motors and much more - this article.
General biology
To begin with, let us recall what kind of organisms they are and what place they occupy in the system of the organic world on our planet. The Bacteria domain unites a huge number of unicellular prokaryotic (without a formed nucleus) organisms.
These living cells appeared on the scene of life almost 4 billion years ago and were the first settlers of the planet. They can be of very different shapes (cocci, rods, vibrios, spirochetes), but most of them are flagellated.
Where do bacteria live? Everywhere. More than 5 × 10 30 live on the planet. There are about 40 million of them in 1 gram of soil, up to 39 trillion live in our body. They can be found at the bottom of the Mariana Trench, in hot "black smokers" at the bottom of the oceans, in the ice of Antarctica, and you currently have up to 10 million bacteria on your hands.
The value is undeniable
Despite their microscopic size (0.5-5 microns), their total biomass on Earth is greater than the biomass of animals and plants combined. Their role in the circulation of substances is irreplaceable, and their properties of consumers (destroyers of organic matter) do not allow the planet to be covered with mountains of corpses.
Well, do not forget about pathogens: the causative agents of plague, smallpox, syphilis, tuberculosis and many other infectious diseases are also bacteria.
Bacteria have found application in human economic activity. Starting from the food industry ( dairy products, cheeses, pickled vegetables, alcoholic drinks), the "green" economy (biofuels and biogas) to the methods of cell engineering and the production of drugs (vaccines, serums, hormones, vitamins).

General morphology
As already mentioned, these unicellular representatives of life do not have a nucleus, their hereditary material (DNA molecules in the form of a ring) are located in a certain area of \u200b\u200bthe cytoplasm (nucleoid). Their cell has a plasma membrane and a dense capsule formed by peptidoglycan murein. Of the cell organelles, bacteria have mitochondria, there may be chloroplasts and other structures with different functions.
Most bacteria are flagella. The tight capsule on the surface of the cell prevents them from moving around by changing the cell itself, as amoeba do. Their flagella are dense protein formations of various lengths and about 20 nm in diameter. Some bacteria have a single flagellum (monotrichous), while others have two (amphitrichous). Sometimes flagella are arranged in bundles (lophotrichous) or cover the entire surface of the cell (peritrichous).
Many of them live as single cells, but some form clusters (pairs, chains, filaments, hyphae).

Movement Features
Flagellated bacteria can move in different ways. Some move only forward, and change direction by tumbling. Some are capable of twitching, others move by sliding.
The flagella of bacteria perform the functions of not only a cellular "oar", but can also be a "boarding" tool.
Until recently, it was believed that the flagellum of a bacterium wags like a snake's tail. Recent studies have shown that the flagellum of bacteria is much more complicated. It works like a turbine. Attached to the drive, it rotates in one direction. The actuator, or flagellar motor of bacteria, is a complex molecular structure that works like a muscle. With the difference that the muscle must relax after contraction, and the bacterial motor works constantly.

The nanomechanism of the flagellum
Without delving into the biochemistry of movement, we note that up to 240 proteins are involved in the creation of the flagellum drive, which are divided into 50 molecular components with a specific function in the system.
In this propulsion system of bacteria, there is a rotor that moves and a stator that provides this movement. There is a drive shaft, bushing, clutch, brakes and accelerators
This miniature engine allows bacteria to travel 35 times their own size in just 1 second. At the same time, the work of the flagellum itself, which makes 60 thousand revolutions per minute, the body spends only 0.1% of all the energy that the cell consumes.
It is also surprising that the bacterium can replace and repair all the spare parts of its motor mechanism “on the go”. Just imagine that you are on an airplane. And technicians change the blades of a running motor.
Flagellate bacterium vs. Darwin
An engine capable of operating at speeds up to 60,000 revolutions per minute, self-starting and using only carbohydrates (sugar) as fuel, having a device akin to an electric motor - could such a device have arisen in the process of evolution?
This is the question that Michael Behe, Ph.D. in biology, asked himself in 1988. He introduced into biology the concept of an irreducible system - a system in which all its parts are simultaneously necessary to ensure its operation, and the removal of at least one part leads to a complete disruption of its functioning.
From the standpoint of Darwin's evolution, all structural changes in the body occur gradually and only successful ones are selected by natural selection.
M. Behe's conclusions, set out in the book "Darwin's Black Box" (1996): the engine of a flagellar bacterium is an indivisible system of more than 40 parts, and the absence of at least one will lead to a complete dysfunction of the system, which means that this system could not have occurred through natural selection .

Balm for creationists
The theory of creation as presented by the scientist and professor of biology, dean of the Faculty of Biological Sciences at the Lehigh University of Bethlehem (USA) M. Behe immediately attracted the attention of church ministers and supporters of the theory of the divine origin of life.
In 2005, in the United States, there was even a lawsuit where Behe was a witness from the supporters of the theory of "reasonable design", which considered the introduction of the study of creationism in the schools of Dover in the course "On pandas and people." The process was lost, the teaching of such a subject was recognized as contrary to the current constitution.
But the debate between creationists and evolutionists continues today.
The flagellum is the surface structure of a bacterial cell, which serves them for movement in liquid media.
Depending on the location of the flagella, bacteria are divided into (Fig. 1):
Peritrichial
mixed
Pole
Subpolar
Pole flagella- one or more flagella are located on one (monopolar) or both (bipolar) poles of the cell and the base is parallel to the long axis of the cell.
Subpolar flagella(subpolar) - one or more flagella are located at the point of transition of the lateral surface to the pole of the cell at one or two of its ends. At the base is a right angle with the long axis of the cell.
Lateral flagella(lateral) - one or more flagella in the form of a bundle are located at the midpoint of one of the halves of the cell.
Peritrichial flagella- located over the entire surface of the cell one by one or in bundles, the poles are usually deprived of them.
Mixed flagella- two or more flagella are located in different points cells.
Depending on the number of flagella, there are:
Monotrichous - one flagellum
Polytrichs - bunch of flagella
Also distinguished:
lophotrichous- monoply polytrichial arrangement of flagella.
amphitriches- bipolar polytrichial arrangement of flagella.
The structure of the bacterial flagellum and basal body. Flagellum.
The flagellum itself is arranged quite simply: a filament that is attached to the basal body. Sometimes a curved section of the tube, the so-called hook, can be inserted between the basal body and the filament; it is thicker than the filament and participates in the flexible attachment of the filament to the basal body.
In terms of chemical composition, the flagellum consists of 98% flagellin protein (flagellum - flagellum), it contains 16 amino acids, glutamine and aspartic acids predominate, tryptophan, cysteine and cystine are absent in a small amount of aromatic amino acids. Flagellin imposes antigenic specificity, it is called the H-antigen. Bacterial flagella do not have ATPase activity.
The thickness of the flagellum is 10–12 nm, the length is 3–15 µm.
It is a rigid spiral, twisted counterclockwise. The rotation of the flagellum is also carried out counterclockwise with a frequency of 40 rpm to 60 rpm, which causes the cell to rotate in the opposite direction, but since the cell is much heavier than the flagellum, then its rotation is slower from 12 to 14 rpm.
The flagellum grows from the distal end, where the subunits enter through the internal channel. In some species, the flagellum is additionally covered on the outside with a sheath, which is a continuation of the cell wall and probably has the same structure.
Basal body
The basal body consists of 4 parts:
Rod mating with filament or hook
Two disks strung on a rod. (M and S)
Group of protein complexes (stators)
protein cap
Bacteria that have an inner and outer membrane have 2 additional disks (P and L) and protein structures that are on the outer membrane near the basal body, hence they do not play an important role in movement.
The peculiarity of the structure of the basal body is determined by the structure of the cell wall: its intactness is necessary for the movement of flagella. Treatment of cells with lysozyme leads to the removal of the peptidoglycan layer from the cell wall, which leads to loss of movement, although the structure of the flagellum was not disturbed.
Bacterial flagella determine the motility of the bacterial cell. Flagella are thin filaments that originate from the cytoplasmic membrane and are longer than the cell itself. The flagella are 12–20 nm thick and 3–15 µm long. They consist of 3 parts: a spiral thread, a hook and a basal body containing a rod with special discs (1 pair of discs for gram-positive and 2 pairs of discs for gram-negative bacteria). The discs of the flagella are attached to the cytoplasmic membrane and cell wall. This creates the effect of an electric motor with a motor rod that rotates the flagellum. Flagella consist of a protein - flagellin (from flagellum - flagellum); is an H antigen. Flagellin subunits are coiled. The number of flagella in bacteria of different species varies from one (monotrich) in Vibrio cholerae to ten or hundreds of flagella extending along the perimeter of the bacterium (peritrich) in Escherichia coli, Proteus, etc. Lofotrichs have a bundle of flagella at one end of the cell. Amphitrichous have one flagellum or a bundle of flagella at opposite ends of the cell.
Pili (fimbriae, villi) - filamentous formations, thinner and shorter (3-10 nm x 0.3-10 microns) than flagella. Pili extend from the cell surface and consist of the pilin protein, which has antigenic activity. There are pili responsible for adhesion, that is, for attaching bacteria to the affected cell, as well as pili responsible for nutrition, water-salt metabolism and sexual (F-pili), or conjugation pili. Drinks are plentiful - several hundred per cage. However, sex pili are usually 1-3 per cell: they are formed by so-called "male" donor cells containing transmissible plasmids (F-, R-, Col-plasmids). A distinctive feature of sex pili is interaction with special "male" spherical bacteriophages, which are intensively adsorbed on sex pili.
Spores are a peculiar form of dormant firmicute bacteria, i.e. bacteria with gram-positive cell wall structure. Spores are formed under unfavorable conditions for the existence of bacteria (drying, nutrient deficiency, etc.. One spore (endospore) is formed inside the bacterial cell. The formation of spores contributes to the preservation of the species and is not a method of reproduction, like in fungi. Spore-forming bacteria of the genus Bacillus have spores that do not exceeding the diameter of the cell.Bacteria whose spore size exceeds the diameter of the cell are called clostridia, for example, bacteria of the genus Clostridium (lat. Clostridium - spindle).The spores are acid-resistant, therefore, they are stained red according to the Aujeszky method or according to the Ziehl-Neelsen method, and the vegetative cell into blue.
The shape of the dispute can be oval, spherical; the location in the cell is terminal, i.e. at the end of the stick (in the causative agent of tetanus), subterminal - closer to the end of the stick (in pathogens of botulism, gas gangrene) and central (in anthrax bacilli). The spore persists for a long time due to the presence of a multi-layered shell, calcium dipicolinate, low water content and sluggish metabolic processes. Under favorable conditions, spores germinate through three successive stages: activation, initiation, germination.
8. The main forms of bacteria
Globular bacteria (cocci) are usually spherical, but may be slightly oval or bean-shaped. Cocci can be located singly (micrococci); in pairs (diplococci); in the form of chains (streptococci) or grape bunches (staphylococci), a package (sarcinas). Streptococci can cause tonsillitis and erysipelas, staphylococci - various inflammatory and purulent processes.
rod-shaped bacteria the most common. Rods can be single, connected in pairs (diplobacteria) or in chains (streptobacteria). The rod-shaped bacteria include Escherichia coli, pathogens of salmonellosis, dysentery, typhoid fever, tuberculosis, etc. Some rod-shaped bacteria have the ability to form under adverse conditions disputes. The spore-forming rods are called bacilli. Spindle-shaped bacilli are called clostridia.
Sporulation is a complex process. Spores differ significantly from a normal bacterial cell. They have a dense shell and a very small amount of water, they do not require nutrients, and reproduction completely stops. Spores are able to withstand drying, high and low temperatures for a long time and can be in a viable state for tens and hundreds of years (spores of anthrax, botulism, tetanus, etc.). Once in a favorable environment, the spores germinate, that is, they turn into the usual vegetative propagating form.
Convoluted bacteria can be in the form of a comma - vibrios, with several curls - spirilla, in the form of a thin twisted stick - spirochetes. Vibrios are the causative agent of cholera, and the causative agent of syphilis is spirochete.
9. Features of the morphology of rickettsia and chlamydia
Rickettsia are small gram-negative microorganisms characterized by pronounced polymorphism - they form cocci, rod-shaped and filamentous forms (Fig. 22). Rickettsia sizes vary from 0.5 to 3-4 microns, the length of filamentous forms reaches 10-40 microns. They do not form spores and capsules, they are stained red according to Zdrodovsky.
Chlamydia are spherical, ovoid or rod-shaped. Their sizes fluctuate within 0.2-1.5 microns. The morphology and size of chlamydia depend on the stage of their intracellular development cycle, which is characterized by the transformation of a small spherical elementary formation into a large initial body with binary division. Before dividing, chlamydia particles are enveloped in a formation resembling a bacterial capsule. Chlamydia stain "* according to Romanovsky-Giemsa, gram-negative, clearly visible in intravital preparations with phase-contrast microscopy.
10. Structure and biology of mycoplasmas.
Only one order, Mycoplasmatales, belongs to the class Mollicutes. Representatives of this order - mycoplasmas -
They differ from bacteria in that they lack a cell wall. Instead, they contain a three-layer lipoprotein cytoplasmic membrane. The sizes of mycoplasmas fluctuate within 125-250 microns. They are round, oval or filamentous, Gram-negative.
Mycoplasmas reproduce by binary fission, like most bacteria, especially after the formation of small coccoid formations (elementary bodies, EB) in filamentous structures.
Mycoplasmas are capable of budding and segmentation. The minimum reproducing unit is ET (0.7-0.2 microns). The main component of the cell membrane is cholesterol. Mycoplasmas are not capable of forming cholesterol and utilize it from tissues or nutrient media supplemented with their introduction. Gram-staining is negative, but Romanovsky-Giemsa staining gives the best results. Mycoplasmas are demanding on cultivation conditions: native serum, cholesterol, nucleic acids, carbohydrates, vitamins and various salts must be added to nutrient media. On dense media, they form characteristic small translucent colonies with a raised granular center, giving them the appearance of "fried eggs". On media with blood, some types of mycoplasmas give a- and beta-hemolysis. In semi-liquid media, mycoplasmas grow along the injection line, forming dispersed, crumbly colonies. In liquid media, they lead to slight haze or opalescence; some strains are able to form the thinnest greasy film. In humans, representatives of the genera Mycoplasma, Ureaplasma and Acholeplasma are isolated, including pathogenic and saprophytic species.