În funcție de locația flagelilor, bacteriile sunt împărțite în. Organele de mișcare: funcții și structură, caracteristici ale mișcării protozoarelor
Structura. Aproximativ jumătate din speciile cunoscute de bacterii de la suprafață au organe de mișcare - flageli curbați ca undă. Masa flagelilor este de până la 2% din masa uscată a bacteriei. Lungimea flagelului este mai mare decât lungimea corpului microorganismului și este de 3–12 μm; grosimea flagelului este de 0,02 μm, iar flagelii polari sunt mai groși decât cei peritric.
Flagelii sunt alcătuiți din proteină flagelină (flagelul latin - flagel), care în structura sa aparține proteinelor contractile de tip miozină. Flagelul conține fie un filament proteic omogen, fie 2-3 filamente strâns pliate într-o împletitură. Firul flagelului este o spirală rigidă răsucită în sens invers acelor de ceasornic; pasul helixului este specific fiecărei specii bacteriene.
Numărul, dimensiunea și aranjamentul flagelilor sunt caracteristici care sunt constante pentru o anumită specie și sunt luate în considerare în taxonomie. Cu toate acestea, unele bacterii pot forma flageli. tipuri diferite. În plus, prezența flagelilor depinde de condițiile de mediu: pe medii solide în timpul cultivării pe termen lung, bacteriile pot pierde flagelii, iar pe medii lichide pot fi dobândite din nou. Numărul și localizarea flagelilor la aceeași specie pot fi determinate de stadiul ciclului de viață. Prin urmare, semnificația taxonomică a acestei caracteristici nu trebuie supraestimată.
Clasificarea bacteriilor în funcție de numărul și localizarea flagelilor:
1. Atrichi - flagelii sunt absenți.
2. Monotrichus- un flagel situat la unul dintre polii celulei (genul Vibrio)- aranjament monopolar monotric al flagelilor, cele mai mobile bacterii.
3. Polytrichus - multe flageli:
– lofotric- mănunchi de flageli la un pol al celulei (naștere Pseudomonas, Burkholderia) - aranjament politric monopolar al flagelilor;
– amfitric- la fiecare pol al celulei se află un mănunchi de flagel (genul Spirillum)- aranjament politricos bipolar al flagelilor;
– peritric- flagelii sunt aranjați fără o ordine anume pe întreaga suprafață a celulei (fam. Enterobacteriaceae(naștere Escherichia, Proteus), fam. bacilaceae, familie Clostidiaceae), numărul de flageli variază de la 6 la 1000 pe celulă, în funcție de tipul de bacterie (Fig. 7).
Fig.7. Opțiuni pentru localizarea flagelilor în bacterii:
1 - monotric, 2 - lophotrich;
3 - amfitric; 4 - peritrich.
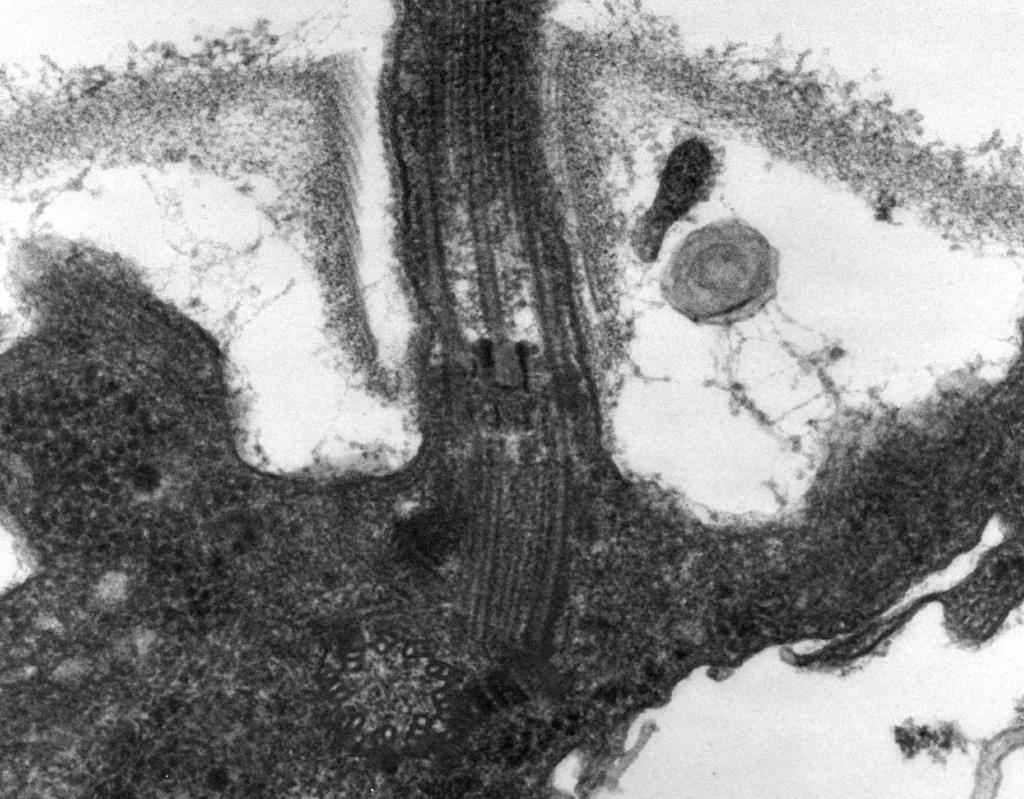
– Microscopia electronică dezvăluie că flagelul este format din trei părți: filament spiralat, cârlig și corp bazal (Fig. 8).
Partea principală a flagelului este lungă fir spiralat (fibrilă) - un cilindru gol, rigid, cu un diametru de aproximativ 120 nm, format din proteină flagelină. Moleculele de proteine formează 11 rânduri pe lungimea firului și sunt dispuse în spirală. În timpul creșterii firului, moleculele de proteine sintetizate în interiorul celulei trec prin cavitatea cilindrului și sunt atașate de o spirală la capătul acesteia. La capatul flagelului se afla un capac proteic care inchide deschiderea cilindrului si previne eliberarea moleculelor de proteine in mediu. Lungimea filamentului flagel poate ajunge la câțiva micrometri. La unele specii de bacterii, flagelul este acoperit suplimentar cu o teacă la exterior. La suprafața CS, firul spirală trece într-o structură curbă îngroșată - un cârlig.

Orez. opt. Schema structurii flagelului
2. Cârlig(20-45 nm grosime) lângă suprafața celulei este un cilindru relativ scurt, compus dintr-o proteină diferită de flagelină și servește la asigurarea unei conexiuni flexibile a filamentului la corpul bazal.
3. Corpul bazal situat la baza flagelului si asigura rotatia acestuia. Corpul bazal conține 9-12 proteine diferite și este format din două sau patru discuri (inele) înșirate pe o tijă, care este o continuare a cârligului. Aceste inele sunt montate în CPM și CS. Cele două inele interioare (M și S) sunt componente obligatorii ale corpului bazal. Inelul M este localizat în CPM, inelul S este situat în spațiul periplasmatic al bacteriilor Gram-negative sau în sacul peptidoglican al bacteriilor Gram-pozitive. Cele două inele exterioare (D și L) sunt opționale pentru mișcare, deoarece sunt prezente numai în bacteriile Gram-negative și sunt localizate, respectiv, în stratul de peptidoglican și în membrana exterioară a CL. Inelele S, D și L sunt imobile și servesc la fixarea flagelului în CS. Rotația flagelului este determinată de rotația inelului M încorporat în CPM-ul celulei. Astfel, caracteristicile structurale ale corpului bazal al flagelului sunt determinate de structura CS.
Din punct de vedere funcțional, corpul bazal este un motor electric alimentat de protoni. Inelul M al corpului bazal (rotorul rotativ) este înconjurat de proteine membranare cu sarcini negative (statorul motor). Celula bacteriană are un mecanism eficient de transformare a energiei electrochimice în energie mecanică. Prin urmare, bacteria cheltuiește aproximativ 0,1% din toată energia pe care o cheltuiește pentru funcționarea flagelului. Atunci când flagelul funcționează, se folosește o forță motrice de protoni, care este oferită de diferența de concentrații de protoni pe părțile exterioare și interioare ale membranei (sunt mai mulți pe partea exterioară) și prezența unei sarcini mai negative. pe partea interioară a membranei. Forța motrice a protonilor forțează protonii să treacă prin corpul bazal în celulă, în timp ce ei rămân în anumite părți ale rotorului, dându-le o sarcină pozitivă, apoi protonii intră în interiorul celulei. Secțiunile încărcate sunt amplasate în așa fel încât să apară o forță atractivă între secțiunile încărcate ale rotorului și stator, inelul M începe să se rotească cu o viteză de aproximativ 300 rpm. Mecanism de rotație: încărcare-reîncărcare a grupului COOH din aminoacizi. Pentru o revoluție completă a inelului, 500-1000 de protoni trebuie să treacă prin corpul bazal. Rotirea inelului M prin axa legată rigid de acesta și cârligul este transmisă filamentului flagel, care funcționează ca o elice sau ca o elice a unei nave. Bacteria înoată atâta timp cât șurubul funcționează, aportul de inerție este extrem de mic.
În plus, bacteriile, chiar și cele moarte, din mediul acvatic se mișcă ca urmare a mișcării browniene. Celula bacteriană este supusă în mod constant impactului moleculelor din jur, care sunt în mișcare termică. Loviturile din diferite direcții aruncă bacteria dintr-o parte în alta.
Tipul de mișcare a flagelului este de rotație. Există două tipuri de mișcare: rectilinie și răsturnată (schimbări aleatorii periodice ale direcției de mișcare). Când flagelii se rotesc în sens invers acelor de ceasornic (aproximativ 1 secundă), la o frecvență de 40-60 rpm (aproape de viteza unui motor electric mediu), firele lor sunt țesute într-un singur mănunchi (Fig. 9a). Rotația flagelilor este transmisă celulei. Deoarece celula este mult mai masivă decât flagelul, începe să se miște în linie dreaptă în direcția opusă, cu o viteză de 3 ori mai mică decât viteza flagelului.
Aceasta asigură mișcarea de translație a celulei, a cărei viteză într-un mediu lichid pentru tipuri diferite bacteriile este de 20–200 μm/s (aceasta corespunde la aproximativ 300–3000 de lungimi ale corpului pe minut) și o mișcare mai lentă pe suprafața mediului solid.
Bacteria poate înota intenționat într-o direcție timp de cel mult 3 secunde, apoi impactul moleculelor din jur o transformă într-o direcție aleatorie. Prin urmare, bacteriile au dezvoltat un mecanism pentru o schimbare spontană a direcției de mișcare - comutarea motorului flagelar. Când începe să se rotească în sensul acelor de ceasornic (aproximativ 0,1 s), bacteria se oprește și se răstoarnă (efectuează o „turburare”) într-o direcție aleatorie. În acest caz, flagelul se împrăștie în direcții diferite (Fig. 9b). În amfitric, atunci când se mișcă, un mănunchi de flageli este întors pe dos (ca o umbrelă întorsă pe dos de vânt). Apoi motorul comută din nou la rotația în sens invers acelor de ceasornic și bacteria înoată din nou în linie dreaptă, dar într-o direcție diferită, aleatorie.
De asemenea, flagelul poate schimba direcția ca răspuns la un stimul extern. Dacă bacteria se deplasează spre concentrația optimă de atractant, flagelul împing celula prin mediu, mișcarea ei rectilinie devine mai lungă, iar frecvența de răsturnare este mai mică, ceea ce îi permite în cele din urmă să se miște în direcția dorită.
Sunt cunoscute cazuri de existență a flagelilor inactivi (paralizați). Pentru mișcarea bacteriilor flagelate, este necesară integritatea (integritatea) CS. Tratamentul celulelor cu lizozim, care duce la îndepărtarea stratului de peptidoglican al CS, determină pierderea capacității bacteriilor de a se mișca, deși flagelii rămân intacți.
Taxiuri de bacterii. Pa mediu inconjurator rămâne neschimbată, bacteriile înoată aleatoriu. Cu toate acestea, mediul este rareori complet omogen. Dacă mediul este eterogen, bacteriile prezintă reacții comportamentale elementare: se mișcă activ în direcția determinată de anumiți factori externi. Astfel de mișcări intenționate determinate genetic ale bacteriilor se numesc taxiuri. În funcție de factor, se disting chemotaxia (un caz special este aerotaxia), fototaxia, magnetotaxia, termotaxia și viscositaxia.
Chemotaxie- mișcarea într-o anumită direcție față de sursa substanței chimice. Substanțe chimiceîmpărțite în două grupe: taxiuri inerte și provocatoare - chemoefectori. Printre chimioefectori se numără substanțele care atrag bacteriile - atracții (zaharuri, aminoacizi, vitamine, nucleotide), și substanțe care le resping - repellente (unii aminoacizi, alcooli, fenoli, ioni anorganici). Atractantul pentru aerobi și repelent pentru procariotele anaerobe este oxigenul molecular. Atractanții sunt adesea reprezentați de substraturi alimentare, deși nu toate substanțele necesare organismului acționează ca atractante. De asemenea, nu toate substanțele otrăvitoare servesc drept repellente și nu toate substanțele otrăvitoare sunt dăunătoare. Astfel, bacteriile nu sunt capabile să reacționeze la niciun compus, ci doar la anumiți și diferiți pentru diferite bacterii.
În structurile de suprafață ale unei celule bacteriene, există molecule speciale de proteine - receptori care se conectează în mod specific la un anumit chemoefector, în timp ce molecula chemoefector nu se modifică, dar în molecula receptor apar modificări conformaționale. Receptorii sunt localizați neuniform pe întreaga suprafață a celulei și sunt concentrați la unul dintre poli. Starea receptorului reflectă concentrația extracelulară a efectorului corespunzător.
Chemotaxia are o valoare adaptativă. De exemplu, formele de vibrio holeric cu chemotaxie afectată sunt mai puțin virulente.
Aerotaxis- Bacteriile care au nevoie de oxigen molecular se acumuleaza in jurul bulelor de aer care au cazut sub o lamela.
Fototaxis- mișcarea către sau departe de lumină, caracteristică bacteriilor fototrofe care folosesc lumina ca sursă de energie.
magnetotaxie- capacitatea bacteriilor acvatice care conțin cristale de minerale care conțin fier de a înota de-a lungul liniilor câmpului magnetic al Pământului.
Termotaxie- mişcarea în direcţia schimbării temperaturii, care are mare importanță pentru unele bacterii patogene.
Viscozitaxis- capacitatea de a răspunde la modificările vâscozității soluției. Bacteriile tind de obicei către un mediu cu o vâscozitate mai mare, ceea ce este de mare importanță pentru speciile patogene.
Bacteriile de alunecare. Capacitatea de a aluneca la o viteză mică (2–11 µm/s) pe un substrat solid sau vâscos a fost găsită la unele procariote, de exemplu, micoplasme.
Există mai multe ipoteze care explică mișcarea de alunecare. Conform ipoteza propulsiei cu reacție se datorează secreției de mucus prin numeroși pori de mucus din SC, drept urmare celula este respinsă din substrat în direcția opusă direcției secreției de mucus. Conform ipoteza undei calatoare mișcarea de alunecare în formele mobile fără flagelați este asociată cu prezența între stratul de peptidoglican și membrana exterioară a CL a unui strat subțire de proteine de fibrile ordonate, similare cu filamentele flagelilor. Mișcarea de rotație a fibrilelor, „lansată” de aceste structuri, duce la apariția unei „unde de călătorie” (proumfări microscopice în mișcare ale CL) pe suprafața celulei, în urma căreia celula este respinsă din substrat. În cele din urmă, structurile care seamănă cu corpurile bazale ale formelor flagelate au fost descrise în unele bacterii glisante.
Funcțiile flagelilor:
1. Asigura aderenta - stadiul inițial proces infecțios.
2. Asigura motilitatea bacteriilor.
3. Determinați specificitatea antigenică, acesta este antigenul H.
Detectarea flagelilor:
1. Microscopia cu contrast de fază a preparatelor native (picături „zdrobite” și „atârnate”). Microscopic, mobilitatea este determinată în celulele culturii zilnice. Pentru a distinge mobilitatea de mișcarea browniană pasivă, la o picătură din cultura studiată se adaugă o picătură dintr-o soluție apoasă 5% de fenol; în acest caz, mișcarea activă se oprește.
2. Microscopia în câmp întunecat a preparatelor native.
3. Microscopia ușoară a preparatelor colorate cu coloranți sau metale. Deoarece flagelii sunt foarte ușor deteriorați în timpul pregătirii preparatului, aceste metode sunt rareori utilizate în practica de zi cu zi.
Flagelii sunt colorați folosind celule crescute pe agar agar. Celulele situate în apropierea apei de condensare sunt selectate cu o buclă bacteriană și transferate cu grijă în apă distilată sterilă la aceeași temperatură cu temperatura de incubare a bacteriilor pe agar oblic, iar bacteriile nu sunt scuturate de pe buclă, ci scufundate cu grijă în apă. Tubul cu bacterii se lasă la temperatura camerei timp de 30 de minute. Se folosește sticlă pură chimic (spălată într-un amestec de crom), pe care se aplică 2-3 picături de suspensie. Suspensia este răspândită pe suprafața de sticlă prin înclinarea ușoară a acesteia. Uscați medicamentul la aer.
Flagelii sunt foarte subțiri, așa că pot fi detectați doar cu o prelucrare specială. Inițial, cu ajutorul gravurii, se realizează umflarea și creșterea dimensiunii lor, iar apoi preparatul este colorat, datorită căruia devin vizibile la microscopie cu lumină.
Mai des folosit metoda de argint conform lui Morozov (Fig. 10):
- preparatul se fixează cu o soluție de acid acetic glacial timp de 1 minut, se spală cu apă;
- se aplica o solutie de tanin (bronzand, facand flagelul mai dens) timp de 1 minut, spalat cu apa;
- preparatul se tratează prin încălzire cu o soluție de impregnare de azotat de argint timp de 1-2 minute, se spală cu apă, se usucă și se microscopează.
Microscopic, sunt vizibile celule maro închis și flageli mai deschisi.

Orez. zece. Identificarea flagelilor prin argintare
Orez. unsprezece. Identificarea flagelilor
prin microscopie electronică
Fig. 4. Microscopia electronică a preparatelor depuse cu metale grele (Fig. 11).
5. Indirect - în funcție de natura creșterii bacteriene atunci când este semănat în agar semi-lichid 0,3%. După incubarea culturilor într-un termostat timp de 1-2 zile, se observă natura creșterii bacteriene:
- în bacterii nemotile (de ex. S.saprofiticul) există o creștere de-a lungul injecției - o „unghie”, iar mediul este transparent;
în bacterii mobile (de ex. E coli) există creștere în partea laterală a injecției, în întreaga coloană de agar - „herringbone”, și turbiditate difuză a mediului.
Toate bacteriileîmpărțit în mobil și imobil. Organele de mișcare ale bacteriilor sunt flageli. Ele sunt formate din flagelină, o proteină care este înrudită structural cu proteinele contractile de tip miozină.
Baza flagelului este un corp bazal, format dintr-un sistem de discuri (blefaroplast: 1 disc - partea exterioară a peretelui celular, 2 discuri - partea interioară a peretelui celular, 3 discuri - membrana citoplasmatică), „încastrate” în citoplasma. membrana si peretele celular. Lungimea flagelului este mai mare decât lungimea corpului microbului însuși.
După numărul de flageliși locația lor, microorganismele mobile sunt împărțite în:
1. Monotric, având un flagel la capătul corpului (cel mai mobil). De exemplu, Vibrio cholerae.
2. Lophotricous, având un mănunchi de flageli la unul dintre polii celulei. De exemplu, Burkholderia (Pseudomonas) pseudomalei este agentul cauzal al melioidozei.
3. Amfitric, având un flagel la ambii poli ai celulei. De exemplu, Spirillum volutans.
4. Peritric, având flageli în jurul întregului perimetru al celulei. De exemplu, Escherichia coli, Salmonella typhi.
Detectarea flagelilor. Flagelii sunt foarte subțiri, așa că pot fi detectați doar cu o prelucrare specială. În special, la început, cu ajutorul unui mordent, se realizează umflarea și creșterea dimensiunii lor, iar apoi preparatul este colorat, datorită căruia devin vizibile la microscopie cu lumină. Flagelii pot fi detectați prin colorare conform Morozov, Leffler, precum și prin microscopie electronică. Flagelii pot fi detectați și prin motilitatea activă a bacteriilor.
Mișcarea microbilor este observată în preparatele de picături „zdrobite” și „atârnate” din culturile vii. Aceste preparate sunt microscopate cu o lentilă uscată sau de imersie într-un câmp întunecat sau în contrast de fază. În plus, motilitatea poate fi determinată de natura creșterii bacteriilor în agar semi-lichid.
A băut din bacterii.
Pili (pili), sinonime: vilozități, fimbrie, sunt filamente subțiri goale de natură proteică care acoperă suprafața celulelor bacteriene. Spre deosebire de flageli, ei nu îndeplinesc o funcție motorie.
Pili se extind de la suprafața celulei și sunt compuse din proteine pilina.
În funcție de scopul lor funcțional, acestea sunt împărțite în 2 tipuri.
1) Pilii de primul tip sunt prezenți în majoritatea bacteriilor, așa că se numesc „vilozități de tip general” (pili comune). Acestea provoacă atașarea sau aderarea bacteriilor la anumite celule ale organismului gazdă. Aderența este etapa inițială a oricărui proces infecțios.
2) Pili de al doilea tip (sinonime: conjugativ, sau sexual - sex pili) sunt disponibile numai în bacteriile donatoare care au o plasmidă specială. Numărul lor este mic - 1-4 pe celulă.
Ferăstrăile pentru podea îndeplinesc următoarele funcții:
1. Participa la transferul de material genetic de la o celulă la alta în timpul conjugării bacteriilor.
2. Adsorb virusuri bacteriene specifice – bacteriofagi
Sporii bacterieni, condițiile de formare, locația, mecanismul și etapele colorării Aujeszky.
controversă- o formă particulară de bacterii în repaus cu un tip de structură a peretelui celular de tip gram-pozitiv.
sporulare- aceasta este o modalitate de conservare a unei specii (genofor) în mediul extern în condiții nefavorabile, și nu o metodă de reproducere.
Se formează sporiîn condiții nefavorabile pentru existența bacteriilor (uscare, deficiență de nutrienți etc.). În interiorul celulei bacteriene se formează un spor (endospor).
Etapele sporulării
1. Pregătitoare. În citoplasma bacteriilor se formează o zonă compactată care nu are apă liberă, numită „zonă sporogenă”, care conține nucleoidul.
2. Stadiul presporilor (prosporilor). În jurul zonei sporogene se formează o teacă a unei membrane citoplasmatice duble.
3. Formarea unui cortex format din peptidoglican și o membrană exterioară cu un conținut ridicat de săruri de calciu și lipide.
4. Stadiul de maturare. Pe partea exterioară a membranei exterioare se formează un înveliș de spori, după care partea vegetativă a celulei este lizată, eliberând sporul.
Dezvoltarea microbiologiei a adus multe descoperiri în ultimele decenii. Și una dintre ele este particularitățile mișcării bacteriilor flagelate. Designul motoarelor acestor organisme antice sa dovedit a fi foarte complex și, conform principiului muncii lor, este foarte diferit de flagelul celor mai apropiate rude ale noastre eucariote ale protozoarelor. Motorul bacteriei flagelate a fost cea mai fierbinte controversă între creaționiști și evoluționiști. Despre bacterii, motoarele lor flagelare și multe altele - acest articol.
Biologie generală
Pentru început, să ne amintim ce fel de organisme sunt și ce loc ocupă ele în sistemul lumii organice de pe planeta noastră. Domeniul Bacterii unește un număr mare de organisme procariote unicelulare (fără nucleu format).
Aceste celule vii au apărut pe scena vieții în urmă cu aproape 4 miliarde de ani și au fost primii coloniști ai planetei. Pot avea forme foarte diferite (coci, bastonașe, vibrioni, spirochete), dar majoritatea sunt flagelate.
Unde trăiesc bacteriile? Pretutindeni. Mai mult de 5 × 10 30 trăiesc pe planetă. Există aproximativ 40 de milioane de ei într-un gram de sol, până la 39 de trilioane trăiesc în corpul nostru. Ele pot fi găsite pe fundul șanțului Marianei, în „fumători negri” fierbinți de pe fundul oceanelor, în gheața Antarcticii, iar în prezent aveți până la 10 milioane de bacterii pe mâini.
Valoarea este de netăgăduit
În ciuda dimensiunii lor microscopice (0,5-5 microni), biomasa lor totală de pe Pământ este mai mare decât biomasa animalelor și plantelor combinate. Rolul lor în circulația substanțelor este de neînlocuit, iar proprietățile lor de consumatori (distrugători de materie organică) nu permit ca planeta să fie acoperită cu munți de cadavre.
Ei bine, nu uitați de agenții patogeni: agenții cauzatori ai ciumei, variolei, sifilisului, tuberculozei și a multor alte boli infecțioase sunt și bacterii.
Bacteriile și-au găsit aplicație în activitatea economică umană. Pornind de la industria alimentară ( lactate, brânzeturi, legume murate, bauturi alcoolice), economia „verde” (biocombustibili și biogaz) la metodele de inginerie celulară și producția de medicamente (vaccinuri, seruri, hormoni, vitamine).

Morfologie generală
După cum sa menționat deja, acești reprezentanți unicelulari ai vieții nu au un nucleu, materialul lor ereditar (moleculele de ADN sub formă de inel) sunt situate într-o anumită zonă a citoplasmei (nucleoid). Celula lor are o membrană plasmatică și o capsulă densă formată din peptidoglican mureină. Dintre organelele celulare, bacteriile au mitocondrii, pot exista cloroplaste și alte structuri cu funcții diferite.
Majoritatea bacteriilor sunt flageli. Capsula strânsă de pe suprafața celulei le împiedică să se miște prin schimbarea celulei în sine, așa cum fac amiba. Flagelii lor sunt formațiuni dense de proteine de diferite lungimi și aproximativ 20 nm în diametru. Unele bacterii au un singur flagel (monotric), în timp ce altele au două (amphitric). Uneori flagelii sunt aranjați în mănunchiuri (lophotricous) sau acoperă întreaga suprafață a celulei (peritrichos).
Multe dintre ele trăiesc ca celule unice, dar unele formează grupuri (perechi, lanțuri, filamente, hife).

Caracteristici de mișcare
Bacteriile flagelate se pot mișca în moduri diferite. Unii se deplasează doar înainte și își schimbă direcția prin răsturnări. Unii sunt capabili să se zvâcnească, alții se mișcă prin alunecare.
Flagelii bacteriilor îndeplinesc funcțiile nu numai de „vâslă” celulară, ci pot fi și un instrument de „încărcare”.
Până nu demult, se credea că flagelul unei bacterii dăinuie ca coada unui șarpe. Studii recente au arătat că flagelul bacteriilor este mult mai complicat. Funcționează ca o turbină. Atașat la unitate, acesta se rotește într-o direcție. Actuatorul sau motorul flagelar al bacteriilor este o structură moleculară complexă care funcționează ca un mușchi. Cu diferența că mușchiul trebuie să se relaxeze după contracție, iar motorul bacterian funcționează constant.

Nanomecanismul flagelului
Fără să ne adâncim în biochimia mișcării, observăm că până la 240 de proteine sunt implicate în crearea unității flagelului, care sunt împărțite în 50 de componente moleculare cu o funcție specifică în sistem.
În acest sistem de propulsie al bacteriilor, există un rotor care se mișcă și un stator care asigură această mișcare. Există un arbore de transmisie, bucșă, ambreiaj, frâne și acceleratoare
Acest motor în miniatură permite bacteriilor să călătorească de 35 de ori propria lor dimensiune în doar 1 secundă. În același timp, prin munca flagelului în sine, care face 60 de mii de rotații pe minut, corpul cheltuiește doar 0,1% din toată energia pe care o consumă celula.
De asemenea, este surprinzător faptul că bacteria poate înlocui și repara toate piesele de schimb ale mecanismului său motor „din mers”. Imaginează-ți doar că ești într-un avion. Și tehnicienii schimbă lamele unui motor în funcțiune.
Bacteria flagelată vs. Darwin
Un motor capabil să funcționeze la viteze de până la 60.000 de rotații pe minut, cu pornire automată și folosind doar carbohidrați (zahăr) drept combustibil, având un dispozitiv asemănător cu un motor electric - ar fi putut apărea un astfel de dispozitiv în procesul de evoluție?
Aceasta este întrebarea pe care și-a pus-o Michael Behe, doctor în biologie, în 1988. El a introdus în biologie conceptul de sistem ireductibil - un sistem în care toate părțile sale sunt simultan necesare pentru a-și asigura funcționarea, iar îndepărtarea a cel puțin unei părți duce la o întrerupere completă a funcționării acestuia.
Din punctul de vedere al evoluției lui Darwin, toate modificările structurale ale corpului apar treptat și numai cele de succes sunt selectate prin selecție naturală.
Concluziile lui M. Behe, expuse în cartea „Darwin’s Black Box” (1996): motorul unei bacterii flagelare este un sistem indivizibil de peste 40 de părți, iar absența a cel puțin uneia va duce la o disfuncție completă a sistem, ceea ce înseamnă că acest sistem nu ar fi putut apărea prin selecție naturală.

Balsam pentru creaționiști
Teoria creației prezentată de savantul și profesorul de biologie, decanul Facultății de Științe Biologice de la Universitatea Lehigh din Bethleem (SUA) M. Behe a atras imediat atenția slujitorilor bisericii și a susținătorilor teoriei divinului originea vieții.
În 2005, în Statele Unite, a existat chiar un proces în care Behe a fost martor din partea susținătorilor teoriei „designului rezonabil”, care a luat în considerare introducerea studiului creaționismului în școlile din Dover în cursul „ Pe panda și oameni.” Procesul s-a pierdut, predarea unei astfel de discipline a fost recunoscută ca fiind contrară actualei constituții.
Dar dezbaterea dintre creationisti si evolutionisti continua si astazi.
Flagelul este structura de suprafață a unei celule bacteriene, care le servește pentru mișcarea în medii lichide.
În funcție de locația flagelului, bacteriile sunt împărțite în (Fig. 1):
Peritrichial
amestecat
Stâlp
Subpolar
Flagelii polari- unul sau mai mulți flageli sunt localizați pe unul (monopolar) sau ambii (bipolar) poli ai celulei și baza este paralelă cu axa lungă a celulei.
Flagelii subpolari(subpolar) - unul sau mai mulți flageli sunt localizați în punctul de tranziție al suprafeței laterale la polul celulei la unul sau două dintre capete. La bază este un unghi drept cu axa lungă a celulei.
Flageli laterali(lateral) - unul sau mai mulți flageli sub formă de mănunchi sunt localizați la mijlocul uneia dintre jumătățile celulei.
Flageli peritrichiali- situati pe toata suprafata celulei unul cate unul sau in fascicule, polii sunt de obicei lipsiti de ei.
Flageli mixte- doi sau mai mulți flageli sunt localizați în puncte diferite celule.
În funcție de numărul de flageli, există:
Monotric - un flagel
Polytrichs - mănunchi de flageli
De asemenea distins:
lofotric- aranjarea monoplă politrihială a flagelilor.
amphitriches- aranjamentul politrihial bipolar al flagelilor.
Structura flagelului bacterian și a corpului bazal. Flagelul.
Flagelul în sine este aranjat destul de simplu: un filament care este atașat de corpul bazal. Uneori, o secțiune curbată a tubului, așa-numitul cârlig, poate fi introdusă între corpul bazal și filament; este mai groasă decât filamentul și participă la atașarea flexibilă a filamentului de corpul bazal.
În ceea ce privește compoziția chimică, flagelul este format din proteină flagelină 98% (flagelul - flagel), conține 16 aminoacizi, predomină glutamina și acizii aspartici, triptofanul, cisteina și cistina sunt absente într-o cantitate mică de aminoacizi aromatici. Flagelina impune specificitate antigenică, se numește antigen H. Flagelii bacterieni nu au activitate ATPaza.
Grosimea flagelului este de 10–12 nm, lungimea este de 3–15 µm.
Este o spirală rigidă, răsucită în sens invers acelor de ceasornic. Rotirea flagelului se efectuează și în sens invers acelor de ceasornic cu o frecvență de la 40 rpm până la 60 rpm, ceea ce face ca celula să se rotească în sens opus, dar deoarece celula este mult mai grea decât flagelul, apoi rotația sa este mai lentă de la 12 la 14 rpm.
Flagelul crește de la capătul distal, unde subunitățile intră prin canalul intern. La unele specii, flagelul este acoperit suplimentar la exterior cu o teacă, care este o continuare a peretelui celular și are probabil aceeași structură.
Corpul bazal
Corpul bazal este format din 4 părți:
Tijă de împerechere cu filament sau cârlig
Două discuri înșirate pe o tijă. (M și S)
Grup de complexe proteice (statori)
capac proteic
Bacteriile care au o membrană interioară și exterioară au 2 discuri suplimentare (P și L) și structuri proteice care se află pe membrana exterioară în apropierea corpului bazal, prin urmare nu joacă un rol important în mișcare.
Particularitatea structurii corpului bazal este determinată de structura peretelui celular: integritatea acestuia este necesară pentru mișcarea flagelilor. Tratamentul celulelor cu lizozim duce la îndepărtarea stratului de peptidoglican din peretele celular, ceea ce duce la pierderea mișcării, deși structura flagelului nu a fost perturbată.
Flagelii bacterieni determină motilitatea celulei bacteriene. Flagelii sunt filamente subțiri care provin din membrana citoplasmatică și sunt mai lungi decât celula însăși. Flagelii au 12–20 nm grosime și 3–15 µm lungime. Sunt formate din 3 părți: un fir spiralat, un cârlig și un corp bazal care conține o tijă cu discuri speciale (1 pereche de discuri pentru bacterii gram-pozitive și 2 perechi de discuri pentru bacterii gram-negative). Discurile flagelilor sunt atașate de membrana citoplasmatică și de peretele celular. Acest lucru creează efectul unui motor electric cu o tijă de motor care rotește flagelul. Flagelii constau dintr-o proteină - flagelină (din flagel - flagel); este un antigen H. Subunitățile flagelinei sunt spiralate. Numărul de flageli în bacterii de diferite specii variază de la unul (monotrich) la Vibrio cholerae la zece sau sute de flageli care se extind de-a lungul perimetrului bacteriei (peritrich) la Escherichia coli, Proteus etc. Lofotrichs au un mănunchi de flageli la unul. capătul celulei. Amphitrichous au un flagel sau un mănunchi de flageli la capetele opuse ale celulei.
Pili (fimbrii, vilozități) - formațiuni filamentoase, mai subțiri și mai scurte (3-10 nm x 0,3-10 microni) decât flagelii. Pili se extind de la suprafața celulei și constau din proteina pilin, care are activitate antigenică. Există pili responsabili de aderență, adică de atașarea bacteriilor la celula afectată, precum și pili responsabili de nutriție, metabolismul apă-sare și pili sexuali (F-pili), sau de conjugare. Băuturile sunt abundente - câteva sute pe cușcă. Cu toate acestea, sex pili sunt de obicei 1-3 per celulă: sunt formați din așa-numitele celule donatoare „masculin” care conțin plasmide transmisibile (F-, R-, Col-plasmide). O trăsătură distinctivă a pili-ului sexual este interacțiunea cu bacteriofagi sferici „masculi” speciali, care sunt absorbiți intens pe pili-ul sexual.
Sporii sunt o formă particulară de bacterii firmicute latente, de exemplu. bacterii cu structura de perete celular gram-pozitiv. Sporii se formează în condiții nefavorabile pentru existența bacteriilor (uscare, deficiență de nutrienți etc.. În interiorul celulei bacteriene se formează un spor (endospor).Formarea sporilor contribuie la conservarea speciei și nu este o metodă de reproducere. , ca la ciuperci. Bacteriile formatoare de spori din genul Bacillus au spori care nu depășesc diametrul celulei.Bacteriile a căror dimensiune sporilor depășește diametrul celulei se numesc clostridii, de exemplu, bacteriile din genul Clostridium (lat. Clostridium - fus). Sporii sunt rezistenți la acizi, prin urmare, sunt colorați în roșu conform metodei Aujeszky sau conform metodei Ziehl-Neelsen, iar celula vegetativă în albastru.
Forma disputei poate fi ovală, sferică; locația în celulă este terminală, adică la capătul bastonului (în agentul cauzator al tetanosului), subterminal - mai aproape de capătul bastonului (la agenții patogeni ai botulismului, gangrena gazoasă) și central (în bacilii antraxului). Sporul persistă mult timp datorită prezenței unei învelișuri multistratificate, dipicolinat de calciu, conținut scăzut de apă și procese metabolice lente. În condiții favorabile, sporii germinează prin trei etape succesive: activare, inițiere, germinare.
8. Principalele forme de bacterii
Bacteriile globulare (coci) sunt de obicei sferice, dar pot fi ușor ovale sau în formă de fasole. Cocii pot fi localizați individual (micrococi); în perechi (diplococi); sub formă de lanțuri (streptococi) sau ciorchini de struguri (stafilococi), un pachet (sarcine). Streptococii pot provoca amigdalită și erizipel, stafilococi - diverse procese inflamatorii și purulente.
bacterii în formă de tijă cel mai comun. Tijele pot fi simple, conectate în perechi (diplobacteria) sau în lanțuri (streptobacterii). Bacteriile în formă de baston includ Escherichia coli, agenți patogeni ai salmonelozei, dizenteriei, febrei tifoide, tuberculozei etc. Unele bacterii în formă de bastonaș au capacitatea de a se forma în condiții nefavorabile. dispute. Tijele formatoare de spori se numesc bacili. Bacili în formă de fus se numesc clostridii.
Sporularea este un proces complex. Sporii diferă semnificativ de o celulă bacteriană normală. Au o coajă densă și o cantitate foarte mică de apă, nu necesită nutrienți, iar reproducerea se oprește complet. Sporii sunt capabili să reziste mult timp la uscare, la temperaturi ridicate și scăzute și pot fi într-o stare viabilă timp de zeci și sute de ani (spori de antrax, botulism, tetanos etc.). Odată ajunși într-un mediu favorabil, sporii germinează, adică se transformă în forma obișnuită de înmulțire vegetativă.
Bacteriile contorte poate fi sub formă de virgulă - vibrios, cu mai multe bucle - spirilla, sub formă de băț subțire răsucit - spirochete. Vibrionii sunt agentul cauzator al holerei, iar agentul cauzal al sifilisului este spirocheta.
9. Caracteristici ale morfologiei rickettsiei și chlamidiei
Rickettsia sunt microorganisme gram-negative mici caracterizate printr-un polimorfism pronunțat - formează coci, forme de baston și filamentoase (Fig. 22). Dimensiunile Rickettsia variază de la 0,5 la 3-4 microni, lungimea formelor filamentoase ajunge la 10-40 microni. Nu formează spori și capsule, ele sunt colorate în roșu conform lui Zdrodovsky.
Chlamydia este sferică, ovoidă sau în formă de tijă. Dimensiunile lor variază între 0,2-1,5 microni. Morfologia și dimensiunea chlamidiei depind de stadiul ciclului lor de dezvoltare intracelulară, care se caracterizează prin transformarea unei mici formațiuni elementare sferice într-un corp inițial mare cu diviziune binară. Înainte de împărțire, particulele de chlamydia sunt învăluite într-o formațiune care seamănă cu o capsulă bacteriană. Colorație de Chlamydia „* conform Romanovsky-Giemsa, gram-negativă, clar vizibilă în preparatele intravitale cu microscopie cu contrast de fază.
10. Structura și biologia micoplasmelor.
Un singur ordin, Mycoplasmatales, aparține clasei Mollicutes. Reprezentanții acestui ordin - micoplasme -
Ele diferă de bacterii prin faptul că le lipsește un perete celular. În schimb, ele conțin o membrană citoplasmatică de lipoproteine cu trei straturi. Dimensiunile micoplasmelor variază între 125-250 microni. Sunt rotunde, ovale sau filamentoase, Gram-negative.
Micoplasmele se reproduc prin fisiune binară, ca majoritatea bacteriilor, mai ales după formarea unor mici formațiuni cocoide (corpi elementari, EB) în structurile filamentoase.
Micoplasmele sunt capabile de înmugurire și segmentare. Unitatea minimă de reproducere este ET (0,7-0,2 microni). Componenta principală a membranei celulare este colesterolul. Micoplasmele nu sunt capabile să formeze colesterol și îl utilizează din țesuturi sau medii nutritive suplimentate cu introducerea lor. Colorația Gram este negativă, dar colorația Romanovsky-Giemsa dă cele mai bune rezultate. Micoplasmele sunt solicitante în condițiile de cultivare: în mediile nutritive trebuie adăugate ser nativ, colesterol, acizi nucleici, carbohidrați, vitamine și diverse săruri. Pe medii dense, formează colonii caracteristice mici translucide cu un centru granular ridicat, dându-le aspectul de „ouă prăjite”. Pe medii cu sânge, unele tipuri de micoplasme dau hemoliză a și beta. În medii semi-lichide, micoplasmele cresc de-a lungul liniei de injectare, formând colonii dispersate, sfărâmicioase. În mediile lichide, acestea conduc la o ușoară ceață sau opalescență; unele tulpini sunt capabile să formeze cea mai subțire peliculă grasă. La om sunt izolați reprezentanți ai genurilor Mycoplasma, Ureaplasma și Acholeplasma, inclusiv speciile patogene și saprofite.