De acordo com a localização dos flagelos, as bactérias são divididas em. Organelas de movimento: funções e estrutura, características do movimento de protozoários
Estrutura. Cerca de metade das espécies conhecidas de bactérias na superfície têm órgãos de movimento - flagelos curvos em forma de onda. A massa dos flagelos é de até 2% da massa seca da bactéria. O comprimento do flagelo é maior que o comprimento do corpo do microrganismo e é de 3 a 12 μm; a espessura do flagelo é de 0,02 μm, e os flagelos polares são mais espessos que os peritríquios.
Os flagelos são compostos pela proteína flagelina (latim flagelo - flagelo), que em sua estrutura pertence a proteínas contráteis do tipo miosina. O flagelo contém um filamento de proteína homogênea ou 2-3 filamentos firmemente dobrados em uma trança. O fio do flagelo é uma espiral rígida torcida no sentido anti-horário; o passo da hélice é específico para cada espécie bacteriana.
O número, tamanho e disposição dos flagelos são características constantes para uma determinada espécie e são levadas em consideração na taxonomia. No entanto, algumas bactérias podem formar flagelos. tipos diferentes. Além disso, a presença de flagelos depende das condições ambientais: em meios sólidos durante o cultivo a longo prazo, as bactérias podem perder flagelos e em meios líquidos podem ser adquiridos novamente. O número e a localização dos flagelos na mesma espécie podem ser determinados pelo estágio do ciclo de vida. Portanto, o significado taxonômico dessa característica não deve ser superestimado.
Classificação das bactérias de acordo com o número e localização dos flagelos:
1. Atrichi - flagelos ausentes.
2. Monótrico- um flagelo localizado em um dos pólos da célula (gênero Vibrio)- arranjo monotríquio monopolar dos flagelos, as bactérias mais móveis.
3. Polítrico - muitos flagelos:
– lofotrico- feixe de flagelos em um pólo da célula (nascimento Pseudomonas, Burkholderia) - arranjo politrico monopolar de flagelos;
– anfítrico- em cada pólo da célula há um feixe de flagelos (gênero Spirillum)- arranjo politrico bipolar dos flagelos;
– peritríquio- flagelos estão dispostos em nenhuma ordem particular sobre toda a superfície da célula (fam. Enterobacteriaceae(nascimento Escherichia, Proteu), fam. bacillaceae, família Clostidiaceae), o número de flagelos varia de 6 a 1000 por célula, dependendo do tipo de bactéria (Fig. 7).
Fig.7. Opções para a localização de flagelos em bactérias:
1 - monotrich, 2 - lophotrich;
3 - anfítrico; 4 - peritrick.
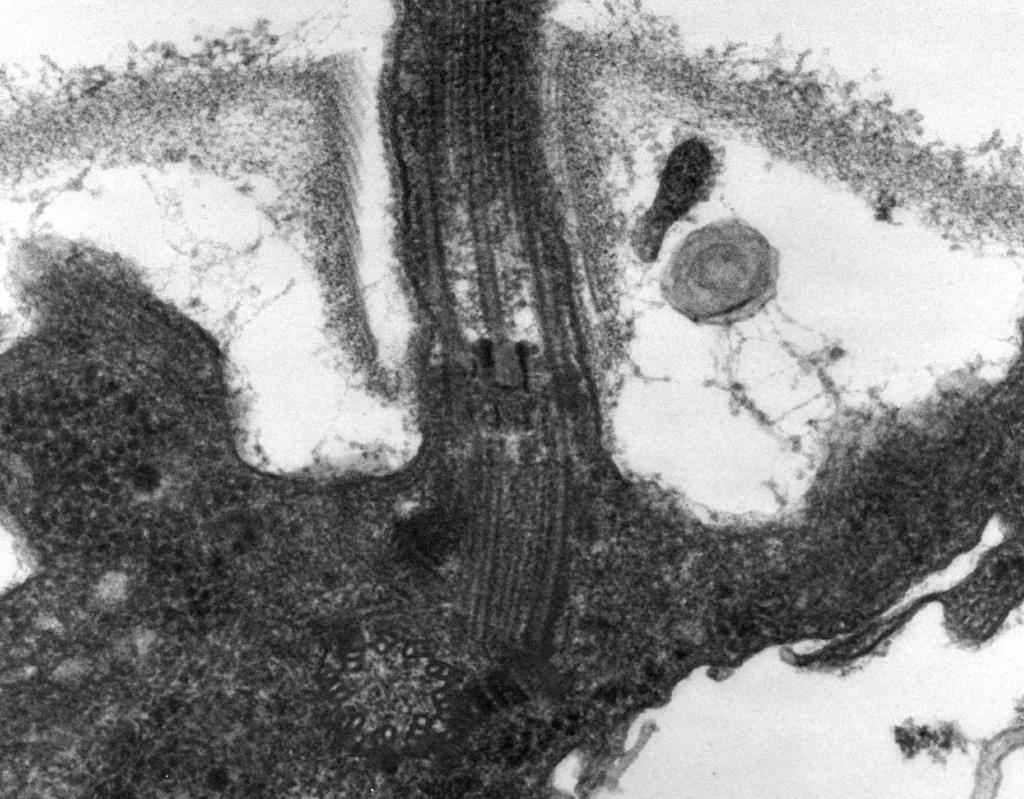
– A microscopia eletrônica revela que flagelo consiste em três partes: filamento espiral, gancho e corpo basal (Fig. 8).
A parte principal do flagelo é longa fio espiral (fibrila) - um cilindro oco rígido com um diâmetro de cerca de 120 nm, constituído por proteína flagelina. As moléculas de proteína formam 11 fileiras ao longo do comprimento do fio e estão dispostas em espiral. Durante o crescimento do fio, moléculas de proteínas sintetizadas no interior da célula passam pela cavidade do cilindro e são presas a uma espiral em sua extremidade. Na extremidade do flagelo existe uma tampa proteica (tampa) que fecha a abertura do cilindro e impede a liberação de moléculas de proteína no ambiente. O comprimento do filamento do flagelo pode atingir vários micrômetros. Em algumas espécies de bactérias, o flagelo é adicionalmente coberto com uma bainha do lado de fora. Na superfície do CS, o fio espiral passa para uma estrutura curvada espessa - um gancho.

Arroz. oito. Esquema da estrutura do flagelo
2. Gancho(20-45 nm de espessura) próximo à superfície da célula é um cilindro relativamente curto, composto por uma proteína diferente da flagelina, e serve para fornecer uma conexão flexível do filamento ao corpo basal.
3. Corpo basal localizado na base do flagelo e garante sua rotação. O corpo basal contém 9 a 12 proteínas diferentes e consiste em dois ou quatro discos (anéis) amarrados em uma haste, que é uma continuação do gancho. Esses anéis são montados no CPM e CS. Os dois anéis internos (M e S) são componentes obrigatórios do corpo basal. O anel M está localizado no CPM, o anel S está localizado no espaço periplasmático de bactérias Gram-negativas ou no saco peptidoglicano de bactérias Gram-positivas. Os dois anéis externos (D e L) são opcionais para o movimento, pois estão presentes apenas em bactérias Gram-negativas e estão localizados, respectivamente, na camada de peptidoglicano e na membrana externa do CL. Os anéis S, D e L são imóveis e servem para fixar o flagelo no SC. A rotação do flagelo é determinada pela rotação do anel M embutido no CPM da célula. Assim, as características estruturais do corpo basal do flagelo são determinadas pela estrutura do SC.
Funcionalmente, o corpo basal é um motor elétrico alimentado por prótons. O anel M do corpo basal (rotor rotativo) é cercado por proteínas de membrana com cargas negativas (estator do motor). A célula bacteriana tem um mecanismo eficiente para converter energia eletroquímica em energia mecânica. Portanto, a bactéria gasta cerca de 0,1% de toda a energia que gasta no funcionamento do flagelo. Quando o flagelo opera, uma força próton-motriz é usada, que é fornecida pela diferença nas concentrações de prótons nos lados externo e interno da membrana (há mais deles no lado externo) e a presença de uma carga mais negativa no lado interno da membrana. A força motriz do próton força os prótons a passarem pelo corpo basal para dentro da célula, enquanto permanecem em certas partes do rotor, dando-lhes uma carga positiva, então os prótons vão para dentro da célula. As seções carregadas estão localizadas de tal forma que uma força atrativa surge entre as seções carregadas do rotor e o estator, o anel M começa a girar a uma velocidade de cerca de 300 rpm. Mecanismo de rotação: carga-recarga do grupo COOH nos aminoácidos. Para uma revolução completa do anel, 500-1000 prótons devem passar pelo corpo basal. A rotação do anel M através do eixo rigidamente conectado a ele e o gancho é transmitido ao filamento do flagelo, que funciona como hélice ou hélice de navio. A bactéria nada enquanto o parafuso estiver funcionando, a contribuição da inércia é extremamente pequena.
Além disso, as bactérias, mesmo mortas, no ambiente aquático se movem como resultado do movimento browniano. A célula bacteriana está constantemente sujeita a impactos das moléculas circundantes, que estão em movimento térmico. Acertos de diferentes direções jogam a bactéria de um lado para o outro.
O tipo de movimento dos flagelos é rotacional. Existem dois tipos de movimento: retilíneo e rolando (mudanças aleatórias periódicas na direção do movimento). Quando os flagelos giram no sentido anti-horário (cerca de 1 segundo), a uma frequência de 40-60 rpm (próximo à velocidade de um motor elétrico médio), seus fios são entrelaçados em um único feixe (Fig. 9a). A rotação dos flagelos é transmitida para a célula. Como a célula é muito mais massiva que o flagelo, ela começa a se mover em linha reta na direção oposta, a uma velocidade 3 vezes menor que a velocidade do flagelo.
Isso garante o movimento de translação da célula, cuja velocidade em um meio líquido para tipos diferentes bactérias é de 20 a 200 μm/s (isso corresponde a cerca de 300 a 3.000 comprimentos de corpo por minuto) e o movimento mais lento na superfície do meio sólido.
A bactéria pode nadar propositalmente em uma direção por não mais que 3 s, então os impactos das moléculas circundantes a transformam em uma direção aleatória. Portanto, as bactérias desenvolveram um mecanismo para uma mudança espontânea na direção do movimento - trocando o motor flagelar. Quando começa a girar no sentido horário (cerca de 0,1 s), a bactéria para e vira (realiza uma “caída”) em uma direção aleatória. Neste caso, os flagelos se espalham em diferentes direções (Fig. 9b). Nos anfítricos, ao se mover, um feixe de flagelos é virado do avesso (como um guarda-chuva virado do avesso pelo vento). Em seguida, o motor muda novamente para a rotação no sentido anti-horário e a bactéria nada novamente em linha reta, mas em uma direção diferente e aleatória.
Os flagelos também podem mudar de direção em resposta a um estímulo externo. Se a bactéria se move em direção à concentração ideal de atrativo, os flagelos empurram a célula através do meio, seu movimento retilíneo se torna mais longo e a frequência de tombamento é menor, o que permite que ela se mova na direção desejada.
São conhecidos casos de existência de flagelos inativos (paralisados). Para o movimento de bactérias flageladas, é necessária a integridade (intacto) do CS. O tratamento das células com lisozima, que leva à remoção da camada de peptidoglicano do SC, causa a perda da capacidade de movimentação da bactéria, embora os flagelos permaneçam intactos.
Táxis de bactérias. Tchau meio Ambiente permanece inalterado, as bactérias nadam aleatoriamente. No entanto, o ambiente raramente é completamente homogêneo. Se o ambiente é heterogêneo, as bactérias exibem reações comportamentais elementares: elas se movem ativamente na direção determinada por certos fatores externos. Esses movimentos intencionais de bactérias geneticamente determinados são chamados de táxis. Dependendo do fator, distinguem-se quimiotaxia (um caso especial é a aerotaxia), fototaxia, magnetotaxia, termotaxia e viscositaxia.
Quimiotaxia- movimento em uma determinada direção em relação à fonte do produto químico. Substancias químicas divididos em dois grupos: táxis inertes e causadores - quimioefetores. Entre os quimioefetores existem substâncias que atraem bactérias - atrativos (açúcares, aminoácidos, vitaminas, nucleotídeos), e substâncias que os repelem - repelentes (alguns aminoácidos, álcoois, fenóis, íons inorgânicos). O atrativo para aeróbios e repelentes para procariontes anaeróbios é o oxigênio molecular. Os atrativos são frequentemente representados por substratos alimentares, embora nem todas as substâncias necessárias para o corpo atuem como atrativos. Além disso, nem todas as substâncias venenosas servem como repelentes e nem todos os repelentes são prejudiciais. Assim, as bactérias não são capazes de reagir a nenhum composto, mas apenas a certos e diferentes para diferentes bactérias.
Nas estruturas de superfície de uma célula bacteriana, existem moléculas especiais de proteínas - receptores que se conectam especificamente a um determinado quimioefetor, enquanto a molécula quimioefetora não muda, mas ocorrem mudanças conformacionais na molécula do receptor. Os receptores estão localizados de forma desigual em toda a superfície da célula e estão concentrados em um dos pólos. O estado do receptor reflete a concentração extracelular do efetor correspondente.
A quimiotaxia tem um valor adaptativo. Por exemplo, formas de cólera vibrio com quimiotaxia prejudicada são menos virulentas.
Aerotaxis- Bactérias que precisam de oxigênio molecular se acumulam em torno de bolhas de ar que caíram sob uma lamela.
Fototaxia- movimento de aproximação ou afastamento da luz, característico de bactérias fototróficas que utilizam a luz como fonte de energia.
magnetotaxia- a capacidade das bactérias aquáticas contendo cristais de minerais contendo ferro de nadar ao longo das linhas do campo magnético da Terra.
Termotaxia- movimento na direção da mudança de temperatura, que tem grande importância para algumas bactérias patogênicas.
Viscositaxia- a capacidade de responder a mudanças na viscosidade da solução. As bactérias geralmente tendem a um meio com maior viscosidade, o que é de grande importância para espécies patogênicas.
Bactérias deslizantes. A capacidade de deslizar em baixa velocidade (2-11 µm/s) sobre um substrato sólido ou viscoso foi encontrada em alguns procariontes, por exemplo, micoplasmas.
Existem várias hipóteses que explicam o movimento de deslizamento. De acordo com hipótese de propulsão a jatoé devido à secreção de muco através de numerosos poros de muco no SC, como resultado, a célula é repelida do substrato na direção oposta à direção da secreção de muco. De acordo com a hipótese da onda viajante O movimento de deslizamento em formas móveis livres de flagelados está associado à presença entre a camada de peptidoglicano e a membrana externa do CL de uma fina camada de proteína de fibrilas ordenadas, semelhantes aos filamentos de flagelos. O movimento rotacional das fibrilas, “lançadas” por essas estruturas, leva ao aparecimento de uma “onda viajante” (protuberâncias microscópicas em movimento do CL) na superfície da célula, fazendo com que a célula seja repelida do substrato. Finalmente, estruturas semelhantes aos corpos basais de formas flageladas foram descritas em algumas bactérias deslizantes.
Funções dos flagelos:
1. Fornecer adesão - Estado inicial processo infeccioso.
2. Fornecer motilidade de bactérias.
3. Determine a especificidade antigênica, este é o antígeno H.
Detecção de flagelos:
1. Microscopia de contraste de fase de preparações nativas (gotas “esmagadas” e “penduradas”). Microscopicamente, a mobilidade é determinada nas células da cultura diária. Para distinguir a mobilidade do movimento browniano passivo, uma gota de uma solução aquosa de fenol a 5% é adicionada a uma gota da cultura estudada; neste caso, o movimento ativo é interrompido.
2. Microscopia de campo escuro de preparações nativas.
3. Microscopia de luz de preparações coradas com corantes ou metais. Uma vez que os flagelos são facilmente danificados durante a preparação da preparação, estes métodos raramente são usados na prática diária.
Os flagelos são corados usando células cultivadas em ágar inclinado. As células localizadas perto da água de condensação são selecionadas com uma alça bacteriana e cuidadosamente transferidas para água destilada estéril na mesma temperatura da temperatura de incubação de bactérias em ágar inclinado, e as bactérias não são retiradas da alça, mas cuidadosamente imersas em água. O tubo com as bactérias é deixado à temperatura ambiente por 30 minutos. É usado vidro quimicamente puro (lavado em uma mistura de cromo), no qual são aplicadas 2-3 gotas de suspensão. A suspensão é espalhada sobre a superfície do vidro inclinando-a suavemente. Seque o medicamento ao ar.
Os flagelos são muito finos, então só podem ser detectados com processamento especial. Inicialmente, com a ajuda do condicionamento, obtém-se o inchaço e o aumento do tamanho e, em seguida, a preparação é colorida, devido à qual se tornam visíveis à microscopia de luz.
Mais frequentemente usado método de prata de acordo com Morozov (Fig. 10):
- a preparação é fixada com uma solução de ácido acético glacial por 1 minuto, lavada com água;
- aplicar uma solução de tanino (curtimento, tornando os flagelos mais densos) por 1 minuto, lavado com água;
- a preparação é tratada por aquecimento com uma solução impregnante de nitrato de prata por 1-2 minutos, lavada com água, seca e microscópica.
Microscopicamente, células marrons escuras e flagelos mais claros são visíveis.

Arroz. dez. Identificação de flagelos por prateação
Arroz. onze. Identificação de flagelos
por microscopia eletrônica
Fig. 4. Microscopia eletrônica de preparações depositadas com metais pesados (Fig. 11).
5. Indiretamente - de acordo com a natureza do crescimento bacteriano quando semeada em ágar semilíquido 0,3%. Após a incubação das culturas em um termostato por 1 a 2 dias, a natureza do crescimento bacteriano é observada:
- em bactérias imóveis (ex. S.saprophyticus) há crescimento ao longo da injeção - uma “unha”, e o meio é transparente;
em bactérias móveis (ex. E. coli) há crescimento ao lado da injeção, em toda a coluna de ágar - "espinha de peixe", e turbidez difusa do meio.
Todas as bactérias dividido em móvel e imóvel. Os órgãos de movimento nas bactérias são os flagelos. Eles são compostos de flagelina, uma proteína que está estruturalmente relacionada às proteínas contráteis do tipo miosina.
Base do flageloé um corpo basal, constituído por um sistema de discos (blefaroplasto: 1 disco - o lado externo da parede celular, 2 discos - o lado interno da parede celular, 3 discos - a membrana citoplasmática), "incorporado" no citoplasma membrana e parede celular. O comprimento do flagelo é maior que o comprimento do corpo do próprio micróbio.
De acordo com o número de flagelos e sua localização, os microrganismos móveis são divididos em:
1. Monótrico, com um flagelo na extremidade do corpo (o mais móvel). Por exemplo, Vibrio cholerae.
2. Lofotríquio, com feixe de flagelos em um dos pólos da célula. Por exemplo, Burkholderia (Pseudomonas) pseudomalei é o agente causador da melioidose.
3. Anfítrico, com flagelo nos dois pólos da célula. Por exemplo, Spirillum volutans.
4. Peritríquio, com flagelos em todo o perímetro da célula. Por exemplo, Escherichia coli, Salmonella typhi.
Detecção de flagelos. Os flagelos são muito finos, então só podem ser detectados com processamento especial. Em particular, a princípio, com a ajuda de um mordente, são alcançados inchaço e aumento em seu tamanho e, em seguida, a preparação é colorida, devido à qual se tornam visíveis à microscopia de luz. Os flagelos podem ser detectados por coloração de acordo com Morozov, Leffler, bem como por microscopia eletrônica. Os flagelos também podem ser detectados pela motilidade ativa das bactérias.
O movimento de micróbios é observado em preparações de gotas "esmagadas" e "penduradas" de culturas vivas. Essas preparações são microscópicas com uma lente seca ou de imersão em campo escuro ou em contraste de fase. Além disso, a motilidade pode ser determinada pela natureza do crescimento das bactérias em ágar semilíquido.
Bebeu de bactérias.
Pili (pili), sinônimos: vilosidades, fímbrias, são finos filamentos ocos de natureza proteica que recobrem a superfície das células bacterianas. Ao contrário dos flagelos, eles não desempenham uma função motora.
Os pili se estendem da superfície celular e são compostos de proteínas pilina.
De acordo com sua finalidade funcional, eles são divididos em 2 tipos.
1) Pili do primeiro tipo estão presentes na maioria das bactérias, por isso são chamados de "vilosidades do tipo geral" (pili comum). Eles causam a fixação ou adesão de bactérias a certas células do organismo hospedeiro. A adesão é o estágio inicial de qualquer processo infeccioso.
2) Pili do segundo tipo (sinônimos: conjugativo ou sexual - pili sexual) estão disponíveis apenas em bactérias doadoras que possuem um plasmídeo especial. Seu número é pequeno - 1-4 por célula.
As serras de piso desempenham as seguintes funções:
1. Participar na transferência de material genético de uma célula para outra durante a conjugação de bactérias.
2. Eles adsorvem vírus bacterianos específicos - bacteriófagos
Esporos bacterianos, condições de formação, localização, mecanismo e estágios da coloração de Aujeszky.
controvérsia- uma forma peculiar de bactérias em repouso com um tipo de estrutura de parede celular gram-positiva.
esporulação- trata-se de uma forma de preservação de uma espécie (genóforo) no ambiente externo em condições adversas, e não de um método de reprodução.
Os esporos são formados em condições desfavoráveis para a existência de bactérias (secagem, deficiência de nutrientes, etc.). Dentro da célula bacteriana, um esporo (endósporo) é formado.
Fases da esporulação
1. Preparatório. No citoplasma das bactérias, forma-se uma área compactada que não possui água livre, chamada de "zona esporógena", que contém o nucleoide.
2. Estágio de presporos (prospores). Ao redor da zona esporógena, uma bainha de uma membrana citoplasmática dupla é formada.
3. Formação de um córtex constituído por peptidoglicano e uma membrana externa com alto teor de sais de cálcio e lipídios.
4. Estágio de maturação. Um envelope de esporos é formado no lado externo da membrana externa, após o que a parte vegetativa da célula é lisada, liberando o esporo.
O desenvolvimento da microbiologia trouxe muitas descobertas nas últimas décadas. E uma delas são as peculiaridades do movimento das bactérias flageladas. O design dos motores desses organismos antigos acabou sendo muito complexo e, de acordo com o princípio de seu trabalho, é muito diferente dos flagelos de nossos parentes eucarióticos mais próximos dos protozoários. O motor da bactéria flagelada tem sido a controvérsia mais quente entre criacionistas e evolucionistas. Sobre bactérias, seus motores flagelares e muito mais - este artigo.
Biologia geral
Para começar, recordemos que tipo de organismos são e que lugar ocupam no sistema do mundo orgânico do nosso planeta. O domínio Bacteria une um grande número de organismos procariontes unicelulares (sem núcleo formado).
Essas células vivas apareceram no cenário da vida há quase 4 bilhões de anos e foram os primeiros colonizadores do planeta. Eles podem ter formas muito diferentes (cocos, bastonetes, vibrios, espiroquetas), mas a maioria deles são flagelados.
Onde vivem as bactérias? Em toda parte. Mais de 5 × 10 30 vivem no planeta. Existem cerca de 40 milhões deles em 1 grama de solo, até 39 trilhões vivem em nosso corpo. Eles podem ser encontrados no fundo da Fossa das Marianas, em "fumadores negros" quentes no fundo dos oceanos, no gelo da Antártida, e atualmente você tem até 10 milhões de bactérias em suas mãos.
O valor é inegável
Apesar de seu tamanho microscópico (0,5-5 mícrons), sua biomassa total na Terra é maior do que a biomassa de animais e plantas combinadas. Seu papel na circulação de substâncias é insubstituível, e suas propriedades de consumidores (destruidores de matéria orgânica) não permitem que o planeta seja coberto por montanhas de cadáveres.
Bem, não se esqueça dos patógenos: os agentes causadores da peste, varíola, sífilis, tuberculose e muitas outras doenças infecciosas também são bactérias.
As bactérias encontraram aplicação na atividade econômica humana. A partir da indústria alimentícia ( lacticínios, queijos, legumes em conserva, bebidas alcoólicas), da economia "verde" (biocombustíveis e biogás) aos métodos de engenharia celular e à produção de medicamentos (vacinas, soros, hormônios, vitaminas).

Morfologia geral
Como já mencionado, esses representantes unicelulares da vida não possuem núcleo, seu material hereditário (moléculas de DNA na forma de um anel) está localizado em uma determinada área do citoplasma (nucleoide). Sua célula possui uma membrana plasmática e uma cápsula densa formada por peptidoglicano mureína. Das organelas celulares, as bactérias possuem mitocôndrias, podendo haver cloroplastos e outras estruturas com diferentes funções.
A maioria das bactérias são flagelos. A cápsula apertada na superfície da célula impede que eles se movam, alterando a própria célula, como fazem as amebas. Seus flagelos são formações proteicas densas de vários comprimentos e cerca de 20 nm de diâmetro. Algumas bactérias têm um único flagelo (monótrico), enquanto outras têm dois (anfítricos). Às vezes, os flagelos estão dispostos em feixes (lofotríquios) ou cobrem toda a superfície da célula (peritríquios).
Muitos deles vivem como células únicas, mas alguns formam aglomerados (pares, cadeias, filamentos, hifas).

Recursos de movimento
As bactérias flageladas podem se mover de diferentes maneiras. Alguns avançam apenas e mudam de direção rolando. Alguns são capazes de se contorcer, outros se movem deslizando.
Os flagelos das bactérias desempenham as funções não apenas de um “remo” celular, mas também podem ser uma ferramenta de “embarque”.
Até recentemente, acreditava-se que o flagelo de uma bactéria balançava como o rabo de uma cobra. Estudos recentes mostraram que o flagelo das bactérias é muito mais complicado. Funciona como uma turbina. Anexado à unidade, ele gira em uma direção. O atuador, ou motor flagelar das bactérias, é uma estrutura molecular complexa que funciona como um músculo. Com a diferença de que o músculo deve relaxar após a contração, e o motor bacteriano trabalha constantemente.

O nanomecanismo do flagelo
Sem nos aprofundarmos na bioquímica do movimento, notamos que até 240 proteínas estão envolvidas na criação do acionamento do flagelo, que são divididos em 50 componentes moleculares com função específica no sistema.
Nesse sistema de propulsão de bactérias, existe um rotor que se movimenta e um estator que proporciona esse movimento. Há um eixo de acionamento, bucha, embreagem, freios e aceleradores
Este motor em miniatura permite que as bactérias viajem 35 vezes seu próprio tamanho em apenas 1 segundo. Ao mesmo tempo, com o trabalho do próprio flagelo, que faz 60 mil rotações por minuto, o corpo gasta apenas 0,1% de toda a energia que a célula consome.
Também é surpreendente que a bactéria possa substituir e reparar todas as peças sobressalentes de seu mecanismo motor “em movimento”. Imagine que você está em um avião. E os técnicos trocam as lâminas de um motor em funcionamento.
Bactéria flagelada vs. Darwin
Um motor capaz de operar a velocidades de até 60.000 rotações por minuto, com partida automática e usando apenas carboidratos (açúcar) como combustível, tendo um dispositivo semelhante a um motor elétrico - tal dispositivo poderia ter surgido no processo de evolução?
Esta é a pergunta que Michael Behe, Ph.D. em biologia, se fez em 1988. Ele introduziu na biologia o conceito de sistema irredutível - um sistema em que todas as suas partes são simultaneamente necessárias para garantir seu funcionamento, e a remoção de pelo menos uma parte leva a uma completa interrupção de seu funcionamento.
Do ponto de vista da evolução de Darwin, todas as mudanças estruturais no corpo ocorrem gradualmente e apenas as bem-sucedidas são selecionadas pela seleção natural.
Conclusões de M. Behe, expostas no livro "Darwin's Black Box" (1996): o motor de uma bactéria flagelar é um sistema indivisível de mais de 40 partes, e a ausência de pelo menos uma levará a uma disfunção completa do sistema, o que significa que este sistema não poderia ter ocorrido através da seleção natural.

Bálsamo para criacionistas
A teoria da criação apresentada pelo cientista e professor de biologia, reitor da Faculdade de Ciências Biológicas da Lehigh University of Bethlehem (EUA) M. Behe atraiu imediatamente a atenção de ministros da igreja e defensores da teoria do divino origem da vida.
Em 2005, nos Estados Unidos, houve até uma ação judicial onde Behe foi testemunha dos adeptos da teoria do "design razoável", que considerava a introdução do estudo do criacionismo nas escolas de Dover no curso " Em pandas e pessoas." O processo foi perdido, o ensino de tal disciplina foi reconhecido como contrário à constituição vigente.
Mas o debate entre criacionistas e evolucionistas continua até hoje.
O flagelo é a estrutura da superfície de uma célula bacteriana, que lhes serve para o movimento em meio líquido.
Dependendo da localização dos flagelos, as bactérias são divididas em (Fig. 1):
Peritriquial
misturado
Pólo
Subpolar
flagelos do pólo- um ou mais flagelos estão localizados em um (monopolar) ou ambos (bipolar) pólos da célula e a base é paralela ao longo eixo da célula.
Flagelos subpolares(subpolar) - um ou mais flagelos estão localizados no ponto de transição da superfície lateral para o pólo da célula em uma ou duas de suas extremidades. Na base é um ângulo reto com o longo eixo da célula.
Flagelos laterais(lateral) - um ou mais flagelos na forma de um feixe estão localizados no ponto médio de uma das metades da célula.
Flagelos peritriquiais- localizados em toda a superfície da célula, um a um ou em feixes, os pólos geralmente são privados deles.
Flagelos mistos- dois ou mais flagelos estão localizados em pontos diferentes células.
Dependendo do número de flagelos, existem:
Monótrico - um flagelo
Polytrichs - bando de flagelos
Distinguem-se também:
lofotrico- arranjo politriquial monopolista dos flagelos.
anfitriões- arranjo politriquial bipolar dos flagelos.
A estrutura do flagelo bacteriano e do corpo basal. Flagelo.
O próprio flagelo é organizado de forma bastante simples: um filamento que está ligado ao corpo basal. Às vezes, uma seção curva do tubo, o chamado gancho, pode ser inserida entre o corpo basal e o filamento; é mais espesso que o filamento e participa da fixação flexível do filamento ao corpo basal.
Em termos de composição química, o flagelo é composto por 98% de proteína flagelina (flagelo - flagelo), contém 16 aminoácidos, predominam glutamina e ácidos aspárticos, triptofano, cisteína e cistina estão ausentes em uma pequena quantidade de aminoácidos aromáticos. A flagelina impõe especificidade antigênica, é chamada de antígeno H. Os flagelos bacterianos não possuem atividade ATPásica.
A espessura do flagelo é de 10 a 12 nm, o comprimento é de 3 a 15 µm.
É uma espiral rígida, torcida no sentido anti-horário. A rotação do flagelo também é realizada no sentido anti-horário com frequência de 40 rpm a 60 rpm, o que faz com que a célula gire no sentido contrário, mas desde a célula é muito mais pesada que o flagelo, então sua rotação é mais lenta de 12 a 14 rpm.
O flagelo cresce a partir da extremidade distal, onde as subunidades entram pelo canal interno. Em algumas espécies, o flagelo é adicionalmente coberto por uma bainha, que é uma continuação da parede celular e provavelmente tem a mesma estrutura.
Corpo basal
O corpo basal consiste em 4 partes:
Acasalamento da haste com filamento ou gancho
Dois discos amarrados em uma haste. (M e S)
Grupo de complexos de proteínas (estators)
tampa de proteína
As bactérias que têm uma membrana interna e externa têm 2 discos adicionais (P e L) e estruturas de proteínas que estão na membrana externa perto do corpo basal, portanto, não desempenham um papel importante no movimento.
A peculiaridade da estrutura do corpo basal é determinada pela estrutura da parede celular: sua integridade é necessária para o movimento dos flagelos. O tratamento das células com lisozima leva à remoção da camada de peptidoglicano da parede celular, o que leva à perda de movimento, embora a estrutura do flagelo não tenha sido perturbada.
Os flagelos bacterianos determinam a motilidade da célula bacteriana. Flagelos são filamentos finos que se originam da membrana citoplasmática e são mais longos que a própria célula. Os flagelos têm 12–20 nm de espessura e 3–15 µm de comprimento. Consistem em 3 partes: um fio espiral, um gancho e um corpo basal contendo uma haste com discos especiais (1 par de discos para bactérias gram-positivas e 2 pares de discos para bactérias gram-negativas). Os discos dos flagelos estão ligados à membrana citoplasmática e à parede celular. Isso cria o efeito de um motor elétrico com uma haste motora que gira o flagelo. Os flagelos consistem em uma proteína - flagelina (de flagelo - flagelo); é um antígeno H. As subunidades de flagelina são enroladas. O número de flagelos em bactérias de diferentes espécies varia de um (monotrico) em Vibrio cholerae a dez ou centenas de flagelos que se estendem ao longo do perímetro da bactéria (peritrich) em Escherichia coli, Proteus, etc. extremidade da célula. Anfítricos têm um flagelo ou um feixe de flagelos nas extremidades opostas da célula.
Pili (fímbrias, vilosidades) - formações filamentosas, mais finas e mais curtas (3-10 nm x 0,3-10 mícrons) que os flagelos. Os pili se estendem da superfície celular e consistem na proteína pilina, que possui atividade antigênica. Existem pili responsáveis pela adesão, ou seja, pela fixação de bactérias à célula afetada, bem como pili responsáveis pela nutrição, metabolismo água-sal e sexual (F-pili), ou pili de conjugação. As bebidas são abundantes - várias centenas por gaiola. No entanto, os pili sexuais são geralmente 1-3 por célula: eles são formados pelas chamadas células doadoras "masculinas" contendo plasmídeos transmissíveis (F-, R-, Col-plasmids). Uma característica distintiva dos pili sexuais é a interação com bacteriófagos esféricos "masculinos" especiais, que são intensamente adsorvidos nos pili sexuais.
Os esporos são uma forma peculiar de bactérias firmes dormentes, ou seja, bactérias com estrutura de parede celular gram-positiva. Os esporos são formados em condições desfavoráveis para a existência de bactérias (secagem, deficiência de nutrientes, etc. , como nos fungos. Bactérias formadoras de esporos do gênero Bacillus possuem esporos que não excedem o diâmetro da célula. Clostridium - fuso).Os esporos são resistentes aos ácidos, portanto, são corados de vermelho de acordo com o método de Aujeszky ou de acordo com o método de Ziehl-Neelsen, e a célula vegetativa em azul.
A forma da disputa pode ser oval, esférica; a localização na célula é terminal, i.e. no final do bastão (no agente causador do tétano), subterminal - mais próximo do final do bastão (em patógenos de botulismo, gangrena gasosa) e central (em bacilos de antraz). O esporo persiste por muito tempo devido à presença de uma concha multicamada, dipicolinato de cálcio, baixo teor de água e processos metabólicos lentos. Sob condições favoráveis, os esporos germinam através de três fases sucessivas: ativação, iniciação, germinação.
8. As principais formas de bactérias
Bactérias globulares (cocos) são geralmente esféricas, mas podem ser ligeiramente ovais ou em forma de feijão. Os cocos podem ser localizados isoladamente (micrococos); em pares (diplococos); na forma de cadeias (estreptococos) ou cachos de uva (estafilococos), um pacote (sarcinas). Os estreptococos podem causar amigdalite e erisipela, estafilococos - vários processos inflamatórios e purulentos.
bactérias em forma de bastonete o mais comum. Os bastonetes podem ser simples, conectados em pares (diplobactérias) ou em cadeias (estreptobactérias). As bactérias em forma de bastonete incluem Escherichia coli, patógenos de salmonelose, disenteria, febre tifóide, tuberculose, etc. Algumas bactérias em forma de bastonete têm a capacidade de se formar sob condições adversas disputas. Os bastonetes formadores de esporos são chamados de bacilos. Os bacilos em forma de fuso são chamados clostrídios.
A esporulação é um processo complexo. Os esporos diferem significativamente de uma célula bacteriana normal. Eles têm uma casca densa e uma quantidade muito pequena de água, não precisam de nutrientes e a reprodução para completamente. Os esporos são capazes de resistir à secagem, altas e baixas temperaturas por muito tempo e podem estar em estado viável por dezenas e centenas de anos (esporos de antraz, botulismo, tétano, etc.). Uma vez em um ambiente favorável, os esporos germinam, ou seja, transformam-se na forma usual de propagação vegetativa.
Bactérias complicadas pode estar na forma de uma vírgula - vibrios, com vários cachos - spirilla, na forma de uma vara fina torcida - espiroquetas. Vibrios são o agente causador da cólera, e o agente causador da sífilis é a espiroqueta.
9. Características da morfologia das riquétsias e clamídias
Rickettsia são pequenos microrganismos gram-negativos caracterizados por polimorfismo acentuado - eles formam cocos, formas em forma de bastonete e filamentosos (Fig. 22). Os tamanhos da Rickettsia variam de 0,5 a 3-4 mícrons, o comprimento das formas filamentosas atinge 10-40 mícrons. Eles não formam esporos e cápsulas, são corados de vermelho de acordo com Zdrodovsky.
As clamídias são esféricas, ovóides ou em forma de bastonete. Seus tamanhos flutuam dentro de 0,2-1,5 mícrons. A morfologia e o tamanho das clamídias dependem do estágio de seu ciclo de desenvolvimento intracelular, caracterizado pela transformação de uma pequena formação elementar esférica em um grande corpo inicial com divisão binária. Antes de se dividirem, as partículas de clamídia são envolvidas em uma formação semelhante a uma cápsula bacteriana. Coloração de clamídia "* de acordo com Romanovsky-Giemsa, gram-negativa, claramente visível em preparações intravitais com microscopia de contraste de fase.
10. Estrutura e biologia dos micoplasmas.
Apenas uma ordem, Mycoplasmatales, pertence à classe Mollicutes. Representantes desta ordem - micoplasmas -
Eles diferem das bactérias por não terem parede celular. Em vez disso, eles contêm uma membrana citoplasmática de lipoproteína de três camadas. Os tamanhos dos micoplasmas flutuam dentro de 125-250 mícrons. São redondos, ovais ou filamentosos, Gram-negativos.
Os micoplasmas se reproduzem por fissão binária, como a maioria das bactérias, principalmente após a formação de pequenas formações cocóides (corpos elementares, EB) em estruturas filamentosas.
Os micoplasmas são capazes de brotamento e segmentação. A unidade de reprodução mínima é ET (0,7-0,2 microns). O principal componente da membrana celular é o colesterol. Os micoplasmas não são capazes de formar colesterol e o utilizam a partir de tecidos ou meios nutrientes suplementados com sua introdução. A coloração de Gram é negativa, mas a coloração de Romanovsky-Giemsa dá os melhores resultados. Os micoplasmas são exigentes nas condições de cultivo: soro nativo, colesterol, ácidos nucléicos, carboidratos, vitaminas e vários sais devem ser adicionados aos meios nutrientes. Em meios densos, formam pequenas colônias translúcidas características com centro granular elevado, dando-lhes a aparência de "ovos fritos". Em meios com sangue, alguns tipos de micoplasmas dão hemólise a e beta. Em meios semilíquidos, os micoplasmas crescem ao longo da linha de injeção, formando colônias dispersas e quebradiças. Em meios líquidos, levam a uma leve turbidez ou opalescência; algumas cepas são capazes de formar o filme gorduroso mais fino. Em humanos, são isolados representantes dos gêneros Mycoplasma, Ureaplasma e Acholeplasma, incluindo espécies patogênicas e saprófitas.