За розташуванням джгутиків бактерії діляться на. Органоїди руху: функції та будова, особливості руху найпростіших
Будова. Близько половини відомих видів бактерій на поверхні мають органи руху – хвилеподібно вигнуті джгутики. Маса джгутиків становить до 2% сухої маси бактерії. Довжина джгутика більша за довжину тіла мікроорганізму і становить 3–12 мкм; товщина джгутика 0,02 мкм, причому полярні джгутики товстіші, ніж перитрихіальні.
Джгутики складаються з білка флагеліну (лат. flagella - джгутик), який за своєю структурою відноситься до скоротливих білків типу міозину. У складі джгутика є або одна гомогенна нитка білкова, або 2-3 нитки, щільно згорнуті в косу. Нитка джгутика – жорстка спіраль, закручена проти годинникової стрілки; крок спіралі специфічний кожному за виду бактерій.
Число, розміри та розташування джгутиків є ознаками, постійними для певного виду, та враховуються при систематиці. Однак у деяких бактерій можуть утворюватися джгутики. різних типів. Крім того, наявність джгутиків залежить від умов довкілля: на твердих середовищах при тривалому культивуванні бактерії можуть втратити джгутики, а на рідких - знову придбати. Кількість та розташування джгутиків в одного і того ж виду може визначатися стадією життєвого циклу. Отже, не варто переоцінювати таксономічне значення цієї ознаки.
Класифікація бактерій за кількістю та розташуванням джгутиків:
1. Атріхи - джгутики відсутні.
2. Монотрихи- один джгутик, розташований на одному з полюсів клітини (рід Vibrio)- монополярне монотрихальне розташування джгутиків, рухливі бактерії.
3. Політрихи - багато джгутиків:
– лофотріхи- пучок джгутиків на одному полюсі клітини (пологи Pseudomonas, Burkholderia) - монополярне політрихальне розташування джгутиків;
– амфітріхи- на кожному полюсі клітини розташовано по пучку джгутику (рід Spirillum)- біполярне політрихальне розташування джгутиків;
– перитріхи- джгутики розташовані без певного порядку по всій поверхні клітини (сім. Enterobacteriaceae(пологи Escherichia, Proteus), сем. Bacillaceae,сім. Clostidiaceae), кількість джгутиків від 6 до 1000 на клітину залежно від виду бактерій (рис. 7).
Рис.7.Варіанти розташування джгутиків у бактерій:
1 – монотрих; 2 – лофотрих;
3 – амфітріх; 4 – перитрих.
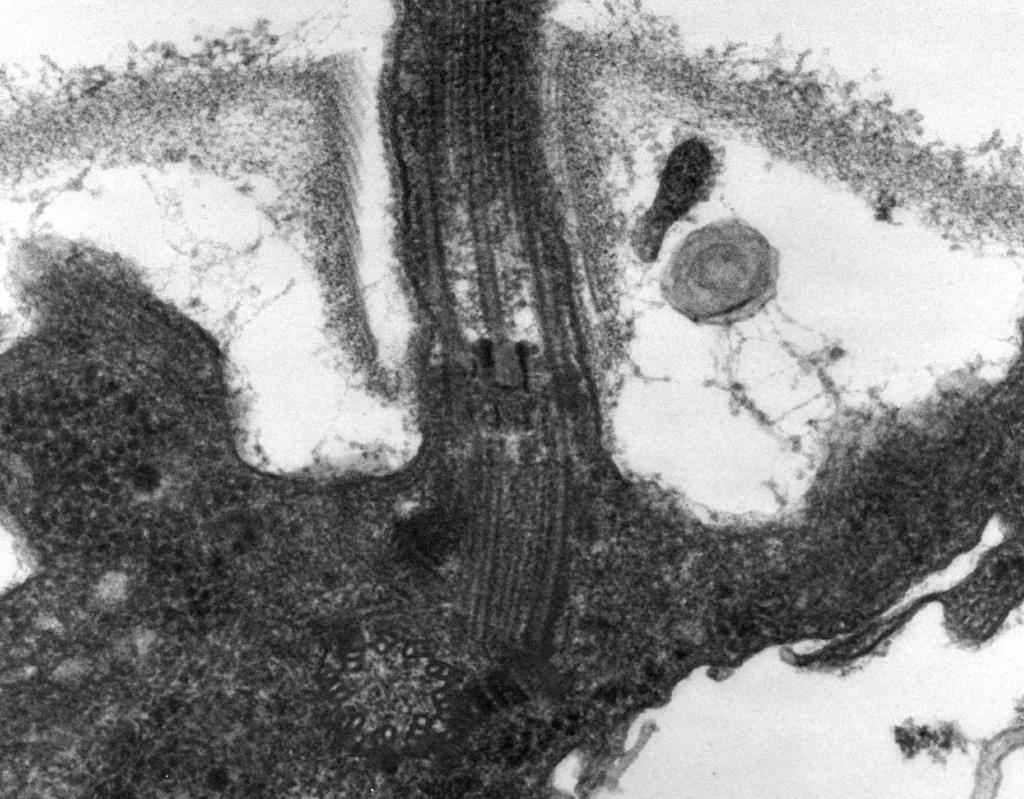
– При електронній мікроскопії виявлено, що джгутик складається з трьох частин:спіральної нитки, гака та базального тіла (рис. 8).
Основну частину джгутика складає довга спіральна нитка (Фібрилла) - жорсткий порожнистий циліндр діаметром близько 120 нм, що складається з білка флагеліну. По довжині нитки білкові молекули утворюють 11 рядів і покладені як спіралі. У процесі зростання нитки білкові молекули, синтезовані всередині клітини, проходять через порожнину циліндра і влаштовуються в спіраль її кінці. На кінці джгутика є білкова шапочка (кришечка), що закриває отвір циліндра і перешкоджає виходу молекул білка в довкілля. Довжина нитки джгутика може досягати кількох мікрометрів. У деяких видів бактерій джгутик зовні додатково вкритий чохлом. У поверхні КС спіральна нитка перетворюється на потовщену вигнуту структуру - гак.

Мал. 8.Схема будови джгутика
2. Гак(Товщиною 20-45 нм) поблизу поверхні клітини - відносно короткий циліндр, складається з білка, що відрізняється від флагеліну, і служить для забезпечення гнучкого з'єднання нитки з базальним тілом.
3. Базальне тілознаходиться на підставі джгутика та забезпечує його обертання. Базальне тіло містить 9-12 різних білків і складається з двох або чотирьох дисків (кілець), нанизаних на стрижень, що є продовженням гака. Ці кільця вмонтовані в ЦПМ та КС. Два внутрішні кільця (M і S) - обов'язкові складові базального тіла. M-кільце локалізоване в ЦПМ, S-кільце розташовується в периплазматичному просторі грамнегативних або в пептидоглікановому мішку грампозитивних бактерій. Два зовнішні кільця (D і L) необов'язкові для руху, так як є тільки у грамнегативних бактерій, локалізовані відповідно в пептидоглікановому шарі і в зовнішній мембрані КС. Кільця S, D і L нерухомі та служать для фіксації джгутика в КС. Обертання джгутика визначається обертанням M-кільця, вбудованого в ЦПМ клітини. Таким чином, особливості будови базального тіла джгутика визначаються будовою КС.
Функціонально базальне тіло є електромотором, що працює на протонах. М-кільце базального тіла (ротор, що обертається), оточене мембранними білками, що мають негативні заряди (статор мотора). Бактеріальна клітина має ефективний механізм, що дозволяє перетворювати електрохімічну енергію на механічну. Тому на роботу джгутика бактерія витрачає близько 0,1% всієї енергії, що нею витрачається. При роботі джгутика використовується протонруха сила, яка забезпечується різницею концентрацій протонів на зовнішній і внутрішній сторонах мембрани (на зовнішній стороні їх більше) і наявністю більш негативного заряду на внутрішній стороні мембрани. Протонруха сила змушує протони проходити через базальне тіло всередину клітини, при цьому вони затримуються на певних ділянках ротора, надаючи їм позитивний заряд, потім протони йдуть всередину клітини. Заряджені ділянки розташовані таким чином, що виникає сила тяжіння між зарядженими ділянками ротора та статора, М-кільце починає обертатися зі швидкістю близько 300 об/c. Механізм обертання: заряджання – перезаряджання групи COOH в амінокислотах. Для повного обігу кільця через базальне тіло має пройти 500-1000 протонів. Обертання М-кільця через жорстко пов'язану з ним вісь і гак передається нитці джгутика, яка функціонує як пропелер або корабельний гвинт. Бактерія пливе до тих пір, поки гвинт працює, внесок інерції виключно малий.
Крім того, бактерії, навіть мертві, що знаходяться у водному середовищі, переміщуються внаслідок броунівського руху. Бактеріальна клітина постійно схильна до ударів навколишніх молекул, що знаходяться в тепловому русі. Удари, що завдаються з різних боків, кидають бактерію з боку на бік.
Тип руху джгутиків – обертальний. Існують два види руху: прямолінійне та перекидання (періодичні випадкові зміни напрямку руху). Коли джгутики обертаються проти ходу годинникової стрілки (близько 1 секунди), з частотою 40-60 об./с (близько швидкості середнього електромотора), їх нитки сплітаються в єдиний джгут (рис. 9а). Обертання джгутиків передається клітині. Так як клітина набагато масивніша за джгутик, вона починає рухатися по прямій у протилежному напрямку, зі швидкістю в 3 рази меншою, ніж швидкість руху джгутика.
Так забезпечується поступальний рух клітини, швидкість якого в рідкому середовищі різних видівбактерій становить 20-200 мкм/с (це відповідає приблизно 300-3000 довжин тіла за хвилину) і повільніше переміщення по поверхні твердих середовищ.
Плити цілеспрямовано в одному напрямку бактерія може не більше 3 с, потім удари навколишніх молекул розгортають її у випадковому напрямку. Тому бактерії виробили механізм спонтанної зміни напрямку руху – перемикання джгутикового двигуна. Коли він починає обертатися протягом годинної стрілки (близько 0,1 с), бактерія зупиняється і перевертається (здійснює «перекид») у випадковому напрямку. При цьому джгутики розлітаються в різні боки (рис. 9б). У амфітріхів при русі один пучок джгутиків вивернуть навиворіт (подібно до парасольки, що вивернула вітром). Потім мотор знову переходить до обертання проти годинникової стрілки, і бактерія знову пливе прямою, але вже в іншому, випадковому напрямку.
Джгутики можуть також змінювати напрямок руху у відповідь на зовнішній стимул. Якщо бактерія переміщається у бік оптимальної концентрації атрактанта, джгутики проштовхують клітину через середовище, її прямолінійний рух стає більш тривалим, а частота перекидів нижчою, що дозволяє їй зрештою переміщатися в потрібному напрямку.
Відомі випадки існування недіючих (паралізованих) джгутиків. Для руху джгутикових бактерій потрібна інтактність (неушкодженість) КС. Обробка клітин лізоцимом, що призводить до видалення пептидогліканового шару КС, викликає втрату здатності бактерій до руху, хоча джгутики залишаються при цьому неушкодженими.
Такси бактерій.Бувай довкіллязалишається незмінною, бактерії плавають безладно. Однак абсолютно однорідне навколишнє середовище буває рідко. Якщо середовище неоднорідне, бактерії виявляють елементарні поведінкові реакції: активно переміщаються у напрямі, що визначається тими чи іншими зовнішніми факторами. Такі генетично детерміновані цілеспрямовані переміщення бактерій називають таксами. Залежно від фактора розрізняють хемотаксис ( окремий випадок- аеротаксис), фототаксис, магнітотаксис, термотаксис та віскозитаксис.
Хемотаксис- рух у певному напрямку щодо джерела хімічної речовини. Хімічні речовиниділять на дві групи: інертні та викликаючі таксиси – хемоефектори. Серед хемоефекторів є речовини, що залучають бактерій, - атрактанти (цукри, амінокислоти, вітаміни, нуклеотиди), та речовини, що їх відлякують, - репеленти (деякі амінокислоти, спирти, феноли, неорганічні іони). Атрактантом для аеробних і репелентом для анаеробних прокаріотів є молекулярний кисень. Атрактанти часто представлені харчовими субстратами, хоча не всі речовини, необхідні для організму, виступають як атрактанти. Також не всі отруйні речовини є репелентами і не всі репеленти шкідливі. Таким чином, бактерії здатні реагувати не на будь-які сполуки, а лише на певні та різні для різних бактерій.
У поверхневих структурах бактеріальної клітини є спеціальні білкові молекули - рецептори, що специфічно з'єднуються з певним хемоефектором, при цьому молекула хемоефектора не змінюється, а в молекулі рецептора відбуваються конформаційні зміни. Рецептори нерівномірно розташовані по всій поверхні клітини, а сконцентровані на одному з полюсів. Стан рецептора відбиває позаклітинну концентрацію відповідного ефектора.
Хемотаксис має пристосувальне значення. Наприклад, форми холерного вібріона з порушеним хемотаксисом виявляються менш вірулентними.
Аеротаксіс- бактерії, що потребують молекулярного кисню, накопичуються навколо бульбашок повітря, що потрапили під покривне скло.
Фототаксис- рух до світла або від нього, властивий фототрофним бактеріям, що використовують світло як джерело енергії.
Магнітотаксис- здатність водних бактерій, що містять кристали залізовмісних мінералів, плисти вздовж ліній магнітного поля Землі.
Термотаксис- рух у бік зміни температури, що має велике значеннядля деяких патогенних бактерій
Віскозитаксис- здатність реагувати зміну в'язкості розчину. Зазвичай бактерії прагнуть у середу з більшою в'язкістю, що має велике значення для патогенних видів.
Ковзання бактерій.Здатність до ковзання з невеликою швидкістю (2-11 мкм/с) за твердим або в'язким субстратом виявлена у деяких прокаріотів, напр., мікоплазм.
Існує кілька гіпотез, що пояснюють ковзний рух. Згідно гіпотезі реактивного рухувоно обумовлено виділенням слизу через численні слизові пори в КС, внаслідок чого клітина відштовхується від субстрату у напрямі, протилежному напрямку виділення слизу. Згідно гіпотезі «біжить хвилі»ковзний рух у рухомих безжгутикових форм пов'язано з наявністю між пептидоглікановим шаром і зовнішньою мембраною КС тонкого білкового шару з упорядковано розташованих фібрил, аналогічних ниткам джгутиків. Обертальний рух фібрил, що «запускається» цими структурами, призводить до появи на поверхні клітини хвилі, що «біжить» (рухаються мікроскопічних опуклостей КС), в результаті чого клітина відштовхується від субстрату. Нарешті, у деяких ковзаючих бактерій описані структури, що нагадують базальні тіла джгутикових форм.
Функції джгутиків:
1. Забезпечують адгезію - початкову стадіюінфекційного процесу.
2. Забезпечують рухливість бактерій.
3. Визначають антигенну специфічність, це Н-антиген.
Виявлення джгутиків:
1. Фазовоконтрастна мікроскопія нативних препаратів («роздавленої» та «висячої» краплі). Мікроскопічно рухливість визначають у клітин добової культури. Для того щоб відрізнити рухливість від пасивного броунівського руху, до краплі досліджуваної культури додають краплю 5% водного розчину фенолу, активний рух в цьому випадку припиняється.
2. Темнопільна мікроскопія нативних препаратів.
3. Світлова мікроскопія пофарбованих барвниками чи металами препаратів. Так як джгутики дуже легко ушкоджуються при приготуванні препарату, у повсякденній практиці ці методи використовують рідко.
Для фарбування джгутиків використовують клітини, вирощені на скошеному агарі. Бактеріальною петлею відбирають клітини, що знаходяться у конденсаційної води і обережно переносять у стерильну дистильовану воду такої ж температури, що і температура інкубування бактерій на скошеному агарі, а бактерії з петлі не струшують, а обережно занурюють у воду. Пробірку з бактеріями залишають за кімнатної температури на 30 хв. Використовують хімічно чисте (вимите у хромовій суміші) скло, на яке наносять 2-3 краплі суспензії. Суспензію розподіляють поверхнею скла, обережно його нахиляючи. Висушують препарат на повітрі.
Джгутики дуже тонкі, тому їх можна виявити тільки при спеціальної обробки. Спочатку за допомогою протруювання досягається розбухання та збільшення їх розміру, а потім проводиться забарвлення препарату, завдяки чому вони стають видимими при світловій мікроскопії.
Найчастіше використовують метод сріблення за Морозовим (рис. 10):
- Препарат фіксують розчином крижаної оцтової кислоти 1 хвилину, промивають водою;
- наносять розчин таніну (дублячий, що робить джгутики більш щільними) на 1 хв, промивають водою;
– обробляють препарат під час підігріву імпрегнуючим розчином азотнокислого срібла 1–2 хв, промивають водою, висушують і мікроскопують.
При мікроскопії видно темно-коричневі клітини та світліші джгутики.

Мал. 10.Виявлення джгутиків методом сріблення
Мал. 11.Виявлення джгутиків
методом електронної мікроскопії
4. Електронна мікроскопія препаратів, напилених важкими металами (рис. 11).
5. Непрямо - за характером зростання бактерій при посіві в напіврідкий 0,3% агар. Після інкубування посівів у термостаті протягом 1–2 діб відзначають характер зростання бактерій:
– у нерухомих бактерій (напр., S. saprophyticus) спостерігається зростання по ходу уколу - «цвях», а середовище прозоре;
– у рухомих бактерій (напр., Е. со1i) спостерігається зростання в сторони від уколу, по всьому стовпчику агару – «ялинка», та дифузне помутніння середовища.
Усі бактеріїподіляються на рухливі та нерухомі. Органами руху у бактерій є джгутики. Вони складаються з білка флагеліну, який за своєю структурою відноситься до скоротливих білків типу міозину.
Підставою джгутикає базальне тільце, що складається із системи дисків (блефаропласту: 1 диск - зовнішня сторона клітинної стінки, 2 диск - внутрішня сторона клітинної стінки, 3 диск - цитоплазматична мембрана), "вмонтованих" у цитоплазматичну мембрану та клітинну стінку. Довжина джгутика більша за довжину тіла самого мікроба.
За кількістю джгутиківта їх розташування рухливі мікроорганізми поділяються на:
1. Монотрихи, що мають на кінці тіла один джгутик (найрухливіші). Наприклад, Vibrio cholerae.
2. Лофотрихи, що мають пучок джгутиків на одному з полюсів клітини. Наприклад, Burkholderia (Pseudomonas) pseudomalei – збудник меліоїдозу.
3. Амфітріхи, які мають джгутик на обох полюсах клітини. Наприклад, Spirillum volutans.
4. Перитрихи, що мають джгутики по всьому периметру клітини. Наприклад, Escherichia coli, Salmonella typhi.
Виявлення джгутиків.Джгутики дуже тонкі, тому їх можна виявити лише при спеціальній обробці. Зокрема, спочатку за допомогою протрави досягається розбухання та збільшення їх розміру, а потім проводиться забарвлення препарату, завдяки чому вони стають видимими при світловій мікроскопії. Виявляти джгутики можна забарвленням за Морозовим, Леффлером, а також електронною мікроскопією. Виявити джгутики можна і за активною рухливістю бактерій.
Рух мікробів спостерігають у препаратах "роздавленої" та "висячої" краплі з живих культур. Мікроскопують ці препарати сухим або іммерсійним об'єктивом у темному полі або у фазовому контрасті. Крім того, рухливість можна визначити за характером зростання бактерій у напіврідкому агарі.
Пили у бактерій.
Пили (pili), синоніми: ворсинки, фімбрії - тонкі порожнисті нитки білкової природи, що покривають поверхню бактеріальних клітин. На відміну від джгутиків не виконують рухової функції.
Пили відходять від поверхні клітини та складаються з білка пиліна.
За своїм функціональним призначенням поділяються на 2 типи.
1) Пили першого типу є у більшості бактерій, тому вони отримали назву "ворсинки загального типу" (common pili). Зумовлюють прикріплення чи адгезію бактерій до певних клітин організму господаря. Адгезія є початковою стадією будь-якого інфекційного процесу.
2) Пили другого типу (синоніми: кон'югативні, або статеві – sex pili) є тільки у бактерій-донорів, що мають спеціальну плазміду. Їхня кількість невелика - 1-4 на клітину.
Статеві пилки виконують такі функції:
1. Беруть участь у передачі генетичного матеріалу від однієї клітини в іншу при кон'югації бактерій.
2. На них адсорбуються специфічні віруси бактерій – бактеріофаги
Суперечки бактерій, умови освіти, розташування, механізм та етапи забарвлення по Ауески.
Спори- своєрідна форма бактерій з грампозитивним типом будови клітинної стінки.
Спороутворення- це спосіб збереження виду (генофора) у зовнішньому середовищі за несприятливих умов, а не спосіб розмноження.
Спори утворюютьсяза несприятливих умов існування бактерій (висушування, дефіцит поживних речовин та ін.). Усередині бактеріальної клітини утворюється одна суперечка (ендоспора).
Стадії спороутворення
1. Підготовча. У цитоплазмі бактерій утворюється ущільнена ділянка, що не має вільної води, яка називається "спорогенною зоною", в якій міститься нуклеоїд.
2. Стадія передспори (проспори). Навколо спорогенної зони утворюється оболонка з подвійної цитоплазматичної мембрани.
3. Утворення кортексу, що складається з пептидоглікану та зовнішньої мембрани з підвищеним вмістом солей кальцію та ліпідів.
4. Стадія дозрівання. Із зовнішньої сторони зовнішньої мембрани утворюється оболонка суперечки, після чого вегетативна частина клітини лізується, звільняючи суперечку.
Розвиток мікробіології приніс в останні десятиліття безліч відкриттів. І одне з них – це особливості руху джгутикових бактерій. Влаштування двигунів цих найдавніших організмів виявилося дуже складним і за принципом своєї роботи дуже відрізняється від джгутиків наших найближчих еукаріотичних родичів найпростіших. Навколо двигуна джгутикової бактерії розгорілися найспекотніші суперечки між креаціоністами та еволюціоністами. Про бактерії, їх джгутикові мотори і багато іншого - дана стаття.
Загальна біологія
Спершу згадаємо, що це за організми і яке місце вони посідають у системі органічного світу на нашій планеті. Домен Bacteria поєднує безліч одноклітинних прокаріотичних (без оформленого ядра) організмів.
Ці живі клітини з'явилися на арені життя майже 4 мільярди років тому і були першими поселенцями планети. Вони можуть бути різної форми (коки, палички, вібріони, спірохети), але більшість з них - джгутикові.
Де мешкають бактерії? Скрізь. На планеті їх мешкає понад 5×10 30 . У 1 г грунту їх налічується близько 40 мільйонів, в нашому організмі мешкає до 39 трильйонів. Їх можна зустріти на дні Маріанської западини, у гарячих «чорних курцях» на дні океанів, у льодах Антарктиди, а на ваших руках зараз перебуває до 10 мільйонів бактерій.
Значення незаперечне
Незважаючи на мікроскопічні розміри (0,5-5 мкм) їхня загальна біомаса на Землі більша, ніж біомаса тварин і рослин, разом узятих. Їх роль кругообігу речовин незамінна, які властивості консументів (руйнівників органіки) неможливо планеті покритися горами трупів.
Ну і не варто забувати про патогени: збудниками чуми, віспи, сифілісу, туберкульозу та багатьох інших інфекційних захворювань теж є бактерії.
Бактерії знайшли застосування і в господарської діяльностілюдини. Починаючи від харчової промисловості ( кисломолочні продукти, сири, квашені овочі, алкогольні напої), «зеленій» економіці (біопаливо та біогаз) до методів клітинної інженерії та виробництва лікарських препаратів (вакцини, сироватки, гормони, вітаміни).

Загальна морфологія
Як уже говорилося, у цих одноклітинних представників життя немає ядра, їхній спадковий матеріал (молекули ДНК у вигляді кільця) розташовані у певній ділянці цитоплазми (нуклеоїд). Клітина їх має плазматичну мембрану та щільну капсулу, утворену пептидогліканом муреїном. З клітинних органел у бактерій є мітохондрії, можуть бути хлоропласти та інші структури з різними функціями.
Більшість бактерій – джгутикові. Щільна капсула на поверхні клітини не дозволяє їм пересуватися за допомогою зміни самої клітини, як це роблять амеби. Їхні джгутики - це щільні білкові утворення різної довжини і діаметром близько 20 нм. Одні бактерії мають єдиний джгутик (монотрихи), в інших їх два (амфітріхи). Іноді джгутики розташовані пучками (лофотріхи) або покривають всю поверхню клітини (перитрихи).
Багато хто з них живе у вигляді одиночних клітин, але деякі утворюють скупчення (пари, ланцюжки, філаменти, гіфи).

Особливості руху
Джгутикові бактерії можуть рухатися по-різному. Одні рухаються тільки вперед, а напрямок змінюють шляхом перекидання. Деякі здатні до посмикування, інші пересуваються шляхом ковзання.
Джгутики бактерій виконують функції як клітинного «весла», але може бути і «абордажным» знаряддям.
Зовсім донедавна вважали, що джгутик бактерії виляє як хвіст у змії. Останні дослідження показали, що джгутик бактерій влаштований значно складніше. Він працює як турбіна. Приєднаний до приводу він обертається в одному напрямку. Привід або джгутиковий мотор бактерії - це складна молекулярна структура, яка працює як м'яз. З тією відмінністю, що м'яз після стиснення повинен розслаблятися, а мотор бактерії працює постійно.

Наномеханізм роботи джгутика
Не заглиблюючись у біохімію руху, відзначимо, що у створенні приводу джгутика долучиться до 240 білків, які діляться на 50 молекулярних складових з певною функцією в системі.
У цій руховій системі бактерії є ротор, що рухається, та статор, який забезпечує цей рух. Є приводний вал, втулка, зчеплення, гальма та прискорювачі
Цей мініатюрний двигун дозволяє бактерії здійснювати переміщення на відстані в 35 разів більші, ніж її власний розмір лише за 1 секунду. При цьому на роботу самого джгутика, що здійснює 60 тисяч обертів за хвилину, організм витрачає всього 0,1% усієї енергії, яку витрачає клітина.
Дивно ще й те, що всі запчастини свого рухового механізму бактерія може замінювати та ремонтувати «на ходу». Просто уявіть, що ви летить у літаку. А техніки змінюють лопаті працюючого двигуна.
Жгутикова бактерія проти Дарвіна
Двигун, здатний працювати зі швидкістю до 60 000 оборотів в хвилину, що самочиниться і використовує у вигляді палива всього лише вуглеводи (цукри), що має пристрій схожий на електричний двигун - чи міг такий пристрій виникнути в процесі еволюції?
Саме це питання поставив собі 1988 року доктор біологічних наук Майкл Біхі. Він увів у біологію поняття нескоротної системи - системи, у якій її частини одночасно необхідні забезпечення її роботи, а видалення бодай однієї частини призводить до повного порушення її функціонування.
З позиції еволюції Дарвіна всі структурні зміни в організмі відбуваються поступово і відбираються природним відбором тільки успішні.
Висновки М. Біхі, викладені у книзі «Чорний ящик Дарвіна» (1996): двигун джгутикової бактерії - неподільна система з більш ніж 40 частин, і відсутність бодай однієї призведе до повної дисфункції системи, а отже, ця система не могла статися шляхом природного відбору .

Бальзам для креаціоністів
Теорія створення у викладі вченого та професора біології, декана факультету біологічних наук у Ліхайському університеті Бетлехема (США) М. Біхі одночасно привернула увагу служителів церкви та прихильників теорії божественного походження життя.
У 2005 році в США навіть відбувся судовий процес, де Біхі виступав свідком з боку прихильників теорії «розумного дизайну», на якому розглядалося питання запровадження в школах Дувру вивчення креаціонізму на курсі «Про пандів і людей». Процес був програний, викладання такого предмета визнали таким, що суперечить чинній конституції.
Але суперечки креаціоністів та еволюціоністів продовжуються і сьогодні.
Джгутик – це поверхнева структура бактеріальної клітини, яка служить їм для руху у рідких середовищах.
Залежно від розташування джгутиків, бактерії поділяються на (Рис.1):
Перитріхіальні
Змішані
Полюсні
Підполюсні
Полюсні джгутики– один або більше джгутиків розташовані на одному (монополярно) або обох (біполярно) полюсах клітини та основа паралельно довгій осі клітини.
Підполюсні джгутики(Субполярні) – один або більше джгутиків розташовані в місці переходу бічної поверхні в полюс клітини на одному або двох її кінцях. В основі - прямий кут з довгою віссю клітини.
Бічні джгутики(Латеральні) - один або більше джгутиків у вигляді пучка розташовані в середній точці однієї з половин клітини.
Перитрихіальні джгутики- Розташовані по всій поверхні клітини по одному або пучками, полюси зазвичай їх позбавлені.
Змішані джгутики– два або кілька джгутиків розташовані в різних точкахклітини.
Залежно від кількості джгутиків, розрізняють:
Монотрихи – один джгутик
Політрихи – пучок джгутиків
Також виділяють:
Лофотріхи– моноплярне політрихіальне розташування джгутиків.
Амфітріхи– біполярне політрихіальне розташування джгутиків.
Будова бактеріального джгутика та базального тіла. Джгутик.
Сам по собі джгутик улаштований досить просто: філамент, що кріпиться до базального тіла. Іноді між базальним тілом і філаментом може бути вставлений вигнутий ділянку трубки, так званий гак, він товщий за філамент, бере участь у гнучкому приєднанні філаменту до базального тіла.
За хімічним складом джгутик на 98% складається з білка флагеліну (flagellum – джгутик), він містить 16 амінокислот, переважають глутамінова та аспарагінова, незначна кількість ароматичних амінокислот відсутні триптофан, цистеїн та цистин. Флагеллін обкладає антигенною специфічністю, його називають Н-антиген. Джгутики бактерій не мають АТФазної активності.
Товщина джгутика 10 - 12 нм, довжина 3-15 мкм.
Є жорсткою спіралью, закрученою проти годинникової стрілки. Обертання джгутика здійснюється також проти годинникової стрілки з частотою від 40 об/сек до 60 об/сек, що викликає обертання клітини протилежному напрямку, але т.к. клітина набагато важча за джгутик, то її обертання повільніше від 12 до 14 об/сек.
Нарощування джгутика відбувається з дистального кінця, куди субодиниці надходять внутрішнім каналом. У деяких видів джгутик зовні додатково покритий чохлом, який є продовженням клітинної стінки і, ймовірно, має таку саму будову.
Базальне тіло
Базальне тіло складається з 4 деталей:
Стрижень, що стикується з філаментом або гаком
Два диски нанизані на стрижень. (М та S)
Група білкових комплексів (статори)
Білковий ковпачок
Бактерії, які мають внутрішню та зовнішню мембрану мають 2 додаткові диски (P та L) та білкові структури, які знаходяться на зовнішній мембрані поблизу базального тіла, отже вони не відіграють важливої ролі у русі.
Особливість будови базального тіла визначається будовою клітинної стінки: її інтактність необхідна для руху джгутиків. Обробка клітин лізоцимом призводить до видалення пептидогліканового шару з клітинної стінки, що веде до втрати руху, хоча будова джгутика порушено не було.
Джгутики бактерій визначають рухливість бактеріальної клітини. Джгутики є тонкими нитками, що беруть початок від цитоплазматичної мембрани, мають більшу довжину, ніж сама клітина. Товщина джгутиків 12-20 нм, довжина 3-15 мкм. Вони складаються з 3 частин: спіралеподібної нитки, гака та базального тільця, що містить стрижень зі спеціальними дисками (1 пара дисків – у грампозитивних та 2 пари дисків – у грамнегативних бактерій). Дисками джгутики прикріплені до цитоплазматичної мембрани та клітинної стінки. При цьому створюється ефект електромотора зі стрижнем-мотором, що обертає джгутик. Джгутики складаються з білка - флагеліну (від flagellum - джгутик); є Н-антигеном. Субодиниці флагеліну закручені у вигляді спіралі. Число джгутиків у бактерій різних видів варіює від одного (монотрих) у холерного вібріона до десятка і сотень джгутиків, що відходять по периметру бактерії (перитрих) у кишкової палички, протею та ін. Лофотріхи мають пучок джгутиків на одному з кінців клітини. Амфітріхи мають по одному джгутику або пучку джгутиків на протилежних кінцях клітини.
Пили (фімбрії, ворсинки) - ниткоподібні утворення, більш тонкі та короткі (3-10нм х 0, 3-10мкм), ніж джгутики. Пили відходять від поверхні клітини і складаються з білка піліну, що має антигенну активність. Розрізняють пили, відповідальні за адгезію, тобто за прикріплення бактерій до клітини, що вражається, а також пили, відповідальні за харчування, водносольовий обмін і статеві (F-пили), або кон'югаційні пили. Пили численні – кілька сотень на клітку. Однак, статевих пилок зазвичай буває 1-3 на клітину: вони утворюються так званими "чоловічими" клітинами-донорами, що містять трансмісивні плазміди (F-, R-, Col-плазміди). Відмінною особливістю статевих пилок є взаємодія з особливими "чоловічими" сферичними бактеріофагами, які інтенсивно адсорбуються на статевих пилях.
Суперечки - своєрідна форма фірмикутних бактерій, що покояться, тобто. бактерій із грампозитивним типом будови клітинної стінки. Спори утворюються при несприятливих умовах існування бактерій (висушування, дефіцит поживних речовин та ін. Усередині бактеріальної клітини утворюється одна суперечка (ендоспора). Утворення суперечки сприяє збереженню виду і не є способом розмноження, як у грибів. Спороутворюючі бактерії роду Bacillus мають суперечки, Бактерії, у яких розмір суперечки перевищує діаметр клітини, називаються клостридіями, наприклад, бактерії роду Clostridium (лат. Clostridium - веретено). у синій колір.
Форма спор може бути овальною, кулястою; розташування у клітині -термінальне, тобто. на кінці палички (у збудника правця), субтермінальне - ближче до кінця палички (у збудників ботулієму, газової гангрени) та центральне (у сибірки-бацили). Суперечка довго зберігається через наявність багатошарової оболонки, дипіколінату кальцію, низького вмісту води та млявих процесів метаболізмів. За сприятливих умов суперечки проростають, проходячи три послідовні стадії: активація, ініціація, проростання.
8. Основні форми бактерій
Кулясті бактерії (коки)зазвичай мають форму кулі, але можуть бути трохи овальної або бобовидної форми. Коки можуть розташовуватися поодинці (мікрококи); попарно (диплококи); у вигляді ланцюжків (стрептококи) або виноградних грон (стафілококи), пакетом (сарцини). Стрептококи можуть викликати ангіну і бешихове запалення, стафілококи - різні запальні та гнійні процеси.
Паличкоподібні бактеріїнайбільш розповсюджені. Палички можуть бути одиночними, попарно з'єднуватися (диплобактерії) або в ланцюжка (стрептобактерії). До паличкоподібних відносяться кишкова паличка, збудники сальмонельозу, дизентерії, черевного тифу, туберкульозу та ін. Деякі паличкоподібні бактерії мають здатність при несприятливих умовах утворювати суперечки.Спороутворюючі палички називають бацилами.Бацили, що нагадують формою веретено, називають клостридіями.
Спороутворення є складним процесом. Спори суттєво відрізняються від звичайної бактеріальної клітини. Вони мають щільну оболонку і дуже малу кількість води, їм не потрібні поживні речовини, а розмноження повністю припиняється. Спори здатні тривалий час витримувати висушування, високі та низькі температури і можуть перебувати в життєздатному стані десятки та сотні років (спори сибірки, ботулізму, правця та ін.). Потрапивши у сприятливе середовище, суперечки проростають, тобто перетворюються на звичайну вегетативну форму, що розмножується.
Покручені бактеріїможуть бути у вигляді коми - вібріони, з декількома завитками - спірили, у вигляді тонкої звивистої палички - спірохети. До вібріонів відноситься збудник холери, а збудник сифілісу – спірохета.
9. Особливості морфології рикетсій та хламідій
Ріккетсії являють собою дрібні грамнегативні мікроорганізми, що характеризуються вираженим поліморфізмом-утворюють кокоподібні, паличкоподібні та ниткоподібні форми (рис. 22). Розміри рикетсії варіюють від 0,5 до 3-4 мкм, довжина ниткоподібних форм досягає 10-40 мкм. Суперечка та капсул не утворюють, забарвлюються за Здродовським у червоний колір.
Хламідії мають кулясту, овоїдну або паличкоподібну форму. Їх розміри коливаються не більше 0,2-1,5 мкм. Морфологія та розміри хламідії залежать від стадії їх внутрішньоклітинного циклу розвитку, для якого характерне перетворення невеликої кулястої елементарної освіти на велике ініціальне тільце з бінарним розподілом. Перед розподілом частинки хламідії обволікаються утворенням, що нагадує бактеріальну капсулу. Хламідії фарбуються "* по Романовському-Гімзі, грамнегативні, добре видно в прижиттєвих препаратах при фазово-контрастній мікроскопії.
10. Будова та біологія мікоплазм.
До класу Mollicutes належить лише один порядок Mycoplasmatales. Представники цього порядку - мікоплазми-
відрізняються від бактерій відсутністю клітинної стінки. Замість неї вони містять тришарову ліпопротеїдну цитоплазматичну мембрану. Розміри мікоплазми коливаються в межах 125-250 мкм. Вони мають форму круглих, овальних або ниткоподібних утворень, грамнегативні
Мікоплазми розмножуються бінарним розподілом, подібно до більшості бактерій, особливо після утворення дрібних коккоподібних утворень (елементарні тільця, ЕТ) у ниткоподібних структурах.
Мікоплазми здатні до брунькування та сегментації. Мінімальною одиницею, що репродукується, вважають ЕТ (0,7-0,2 мкм). Основний компонент клітинної мембрани – холестерин. Мікоплазми не здатні до утворення холестерину та утилізують його з тканин або живильних середовищ, доповнених їх внесенням. За Грамом забарвлюються негативно, але найкращі результати дає забарвлення по Романівському-Гімзі. Мікоплазми вимогливі до умов культивування: до живильного середовища необхідно вносити нативну сироватку, холестерин, нуклеїнові кислоти, вуглеводи, вітаміни та різні солі. На щільних середовищах утворюють характерні дрібні напівпрозорі колонії з піднятим зернистим центром, що надає їм вигляду «яєчні-глазуньі». На середовищах із кров'ю деякі види мікоплазм дають а- та бета-гемоліз. У напіврідких середовищах мікоплазми ростуть по лінії уколу, формуючи дисперсні, кришкуваті колонії. У рідких середовищах призводять до незначного помутніння чи опалесценції; деякі штами здатні утворювати найтоншу жирну плівку. У людини виділяють представників пологів Mycoplasma, Ureaplasma та Acholeplasma, що включають патогенні та сапрофітичні види