W zależności od lokalizacji wici bakterie dzielą się na. Organelle ruchu: funkcje i budowa, cechy ruchu pierwotniaków
Struktura. Około połowa znanych gatunków bakterii na powierzchni ma narządy ruchu - przypominające fale zakrzywione wici. Masa wici stanowi do 2% suchej masy bakterii. Długość wici jest dłuższa niż długość ciała mikroorganizmu i wynosi 3–12 μm; grubość wici wynosi 0,02 μm, a wici polarne są grubsze niż wici peritrichiczne.
Wici zbudowane są z białka flageliny (łac. Wić zawiera albo jedno jednorodne włókno białkowe, albo 2–3 włókna ciasno złożone w warkocz. Nić wici jest sztywną spiralą skręconą w kierunku przeciwnym do ruchu wskazówek zegara; skok helisy jest specyficzny dla każdego gatunku bakterii.
Liczba, wielkość i układ wici to cechy stałe dla danego gatunku i uwzględniane w taksonomii. Jednak niektóre bakterie mogą tworzyć wici. różne rodzaje. Ponadto obecność wici zależy od warunków środowiskowych: na podłożach stałych podczas długotrwałej hodowli bakterie mogą utracić wici, a na podłożach płynnych mogą zostać ponownie pozyskane. Liczbę i lokalizację wici u tego samego gatunku można określić na podstawie etapu cyklu życiowego. Nie należy zatem przeceniać znaczenia taksonomicznego tej cechy.
Klasyfikacja bakterii ze względu na liczbę i umiejscowienie wici:
1. Atrichi - brak wici.
2. Monotrych- jedna wici znajdująca się na jednym z biegunów komórki (rodzaj Vibrio)- monopolarny monotrichiczny układ wici, najbardziej ruchliwych bakterii.
3. Polytrichus - wiele wici:
– lofotryczny- wiązka wici na jednym biegunie komórki (narodziny Pseudomonas, Burkholderia) - monopolarny politrychiczny układ wici;
– amfitryczny- na każdym biegunie komórki znajduje się wiązka wici (rodzaj Spirillum)- dwubiegunowy, politrychiczny układ wici;
– peritrichous-wici ułożone są w przypadkowej kolejności na całej powierzchni komórki (fam. Enterobacteriaceae(narodziny Escherichia, Proteusz), fam. pałeczki, rodzina Clostidiaceae), liczba wici waha się od 6 do 1000 na komórkę, w zależności od rodzaju bakterii (ryc. 7).
Ryc.7. Opcje lokalizacji wici u bakterii:
1 - monotrych, 2 - lofotryk;
3 - amfitryczny; 4 - perytrycz.
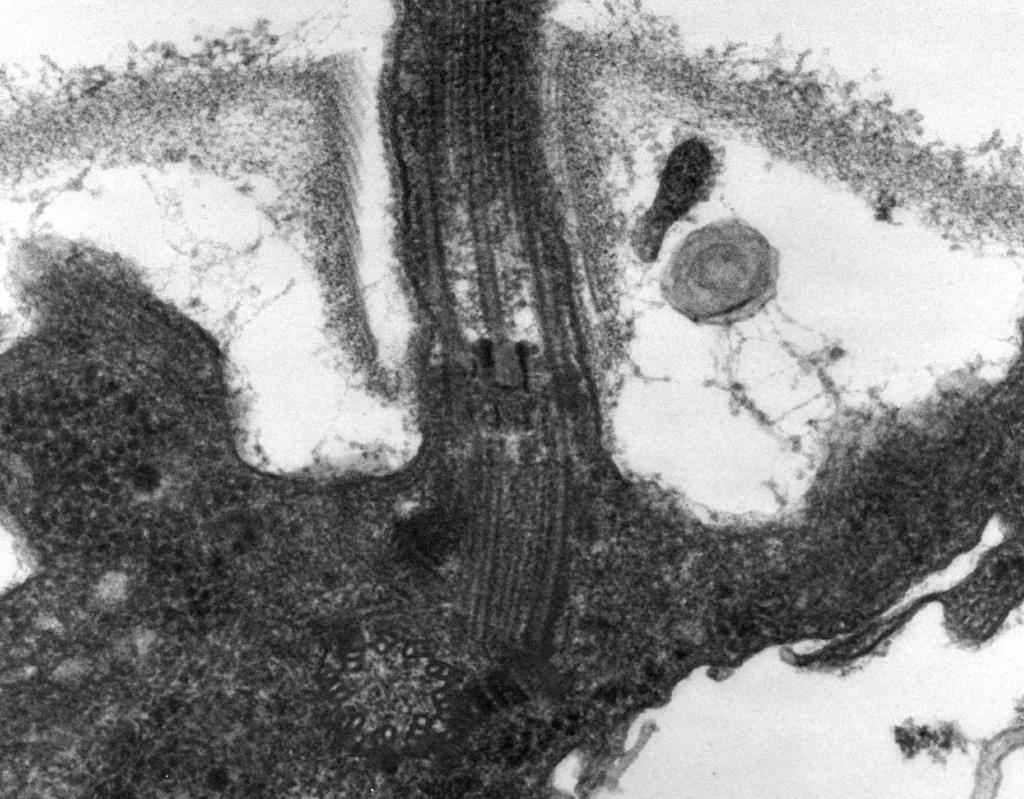
– Pokazuje to mikroskopia elektronowa wić składa się z trzech części: włókno spiralne, haczyk i korpus podstawowy (ryc. 8).
Główna część wici jest długa spiralna nić (fibryla) - sztywny wydrążony cylinder o średnicy około 120 nm, składający się z białka flageliny. Cząsteczki białka tworzą 11 rzędów na całej długości nici i są ułożone spiralnie. Podczas wzrostu nici cząsteczki białka syntetyzowane wewnątrz komórki przechodzą przez wnękę cylindra i przyczepiają się do spirali na jego końcu. Na końcu wici znajduje się białkowa nasadka (wieczko), która zamyka otwór cylindra i zapobiega uwalnianiu cząsteczek białka do środowiska. Długość włókna wici może osiągnąć kilka mikrometrów. U niektórych gatunków bakterii wić jest dodatkowo pokryta osłonką na zewnątrz. Na powierzchni CS spiralna nić przechodzi w pogrubioną zakrzywioną strukturę - haczyk.

Ryż. 8. Schemat budowy wici
2. Hak(o grubości 20–45 nm) w pobliżu powierzchni komórki znajduje się stosunkowo krótki cylinder, składający się z białka innego niż flagelina, który służy do zapewnienia elastycznego połączenia włókna z korpusem podstawowym.
3. Ciało podstawowe znajduje się u podstawy wici i zapewnia jej rotację. Ciało podstawowe zawiera 9–12 różnych białek i składa się z dwóch lub czterech krążków (pierścieni) nawleczonych na pręt, który jest kontynuacją haczyka. Pierścienie te są montowane w CPM i CS. Dwa pierścienie wewnętrzne (M i S) są obowiązkowymi składnikami ciała podstawowego. Pierścień M zlokalizowany jest w CPM, pierścień S w przestrzeni peryplazmatycznej bakterii Gram-ujemnych lub w worku peptydoglikanowym bakterii Gram-dodatnich. Dwa zewnętrzne pierścienie (D i L) są opcjonalne dla ruchu, ponieważ są obecne tylko u bakterii Gram-ujemnych i są zlokalizowane odpowiednio w warstwie peptydoglikanu iw zewnętrznej błonie CL. Pierścienie S, D i L są nieruchome i służą do mocowania wici w CS. Obrót wici jest określony przez obrót pierścienia M wbudowanego w CPM komórki. Zatem cechy strukturalne korpusu podstawowego wici są określone przez strukturę CS.
Funkcjonalnie ciało podstawowe jest silnikiem elektrycznym napędzanym protonami. Pierścień M korpusu podstawowego (wirnik obrotowy) otoczony jest białkami błonowymi o ładunkach ujemnych (stojan silnika). Komórka bakteryjna posiada sprawny mechanizm przekształcania energii elektrochemicznej w energię mechaniczną. Dlatego bakteria wydaje około 0,1% całej energii, którą zużywa na działanie wici. Podczas pracy wici wykorzystywana jest siła protonomotoryczna, którą zapewnia różnica stężeń protonów po zewnętrznej i wewnętrznej stronie błony (jest ich więcej po stronie zewnętrznej) oraz obecność bardziej ujemnego ładunku po wewnętrznej stronie membrany. Siła napędowa protonów zmusza protony do przejścia przez ciało podstawowe do komórki, podczas gdy pozostają one w niektórych częściach wirnika, dając im ładunek dodatni, a następnie protony wchodzą do wnętrza komórki. Naładowane sekcje są rozmieszczone w taki sposób, że między naładowanymi sekcjami wirnika i stojana powstaje siła przyciągania, pierścień M zaczyna się obracać z prędkością około 300 obr./min. Mechanizm rotacji: ładowanie-ładowanie grupy COOH w aminokwasach. Do pełnego obrotu pierścienia przez ciało podstawowe musi przejść 500–1000 protonów. Obrót pierścienia M przez sztywno z nim połączoną oś i hak jest przenoszony na włókno wici, które działa jak śruba napędowa lub śruba napędowa statku. Bakteria pływa tak długo, jak śruba pracuje, wkład bezwładności jest niezwykle mały.
Ponadto bakterie, nawet te martwe, w środowisku wodnym poruszają się w wyniku ruchów Browna. Komórka bakteryjna jest nieustannie poddawana oddziaływaniom otaczających ją cząsteczek, które są w ruchu termicznym. Uderzenia z różnych kierunków rzucają bakterią z boku na bok.
Rodzaj ruchu wici jest obrotowy. Istnieją dwa rodzaje ruchu: prostoliniowy i przewracający (okresowe losowe zmiany kierunku ruchu). Kiedy wici obracają się w kierunku przeciwnym do ruchu wskazówek zegara (około 1 sekundy), z częstotliwością 40–60 obr./min (zbliżoną do prędkości przeciętnego silnika elektrycznego), ich nitki splatają się w jedną wiązkę (ryc. 9a). Rotacja wici jest przekazywana do komórki. Ponieważ komórka jest znacznie masywniejsza niż wici, zaczyna poruszać się po linii prostej w przeciwnym kierunku, z prędkością 3 razy mniejszą niż prędkość wici.
Zapewnia to ruch translacyjny komórki, którego prędkość w płynnym ośrodku dla różne rodzaje bakterii wynosi 20–200 μm/s (odpowiada to około 300–3000 długości ciała na minutę) i wolniejszy ruch na powierzchni podłoża stałego.
Bakteria może celowo płynąć w jednym kierunku przez nie więcej niż 3 s, a następnie uderzenia otaczających cząsteczek obracają ją w losowym kierunku. Dlatego bakterie wykształciły mechanizm spontanicznej zmiany kierunku ruchu – przełączenia motoryki wiciowej. Kiedy zaczyna obracać się zgodnie z ruchem wskazówek zegara (około 0,1 s), bakteria zatrzymuje się i przewraca (wykonuje „przewrót”) w przypadkowym kierunku. W tym przypadku wici rozpraszają się w różnych kierunkach (ryc. 9b). W amphitrichosie podczas ruchu jedna wiązka wici jest wywrócona na lewą stronę (jak parasol wywrócony na lewą stronę przez wiatr). Następnie silnik ponownie przełącza się na obrót przeciwny do ruchu wskazówek zegara, a bakteria ponownie płynie w linii prostej, ale w innym, losowym kierunku.
Wici mogą również zmieniać kierunek w odpowiedzi na bodziec zewnętrzny. Jeśli bakteria porusza się w kierunku optymalnego stężenia atraktantu, wici przepychają komórkę przez pożywkę, jej ruch prostoliniowy staje się dłuższy, a częstotliwość bębnowania jest mniejsza, co ostatecznie pozwala jej poruszać się w pożądanym kierunku.
Znane są przypadki istnienia nieczynnej (sparaliżowanej) wici. Do przemieszczania się bakterii wiciowych niezbędna jest integralność (nienaruszalność) CS. Traktowanie komórek lizozymem, które prowadzi do usunięcia warstwy peptydoglikanowej kortykosteroidów, powoduje utratę zdolności ruchu bakterii, chociaż wici pozostają nienaruszone.
Taksówki bakterii. Do widzenia środowisko pozostaje niezmieniona, bakterie pływają losowo. Jednak środowisko rzadko jest całkowicie jednorodne. Jeśli środowisko jest niejednorodne, bakterie wykazują elementarne reakcje behawioralne: aktywnie poruszają się w kierunku określonym przez określone czynniki zewnętrzne. Takie genetycznie zdeterminowane celowe ruchy bakterii nazywane są taksówkami. W zależności od czynnika wyróżnia się chemotaksję (szczególnym przypadkiem jest aerotaksja), fototaksję, magnetotaksję, termotaksję i wiskozytaksję.
Chemotaksja- ruch w określonym kierunku względem źródła substancji chemicznej. Substancje chemiczne dzieli się na dwie grupy: obojętne i powodujące taksówki - chemoeffektory. Wśród chemoefektorów znajdują się substancje przyciągające bakterie – atraktanty (cukry, aminokwasy, witaminy, nukleotydy) oraz substancje, które je odpychają – repelenty (niektóre aminokwasy, alkohole, fenole, jony nieorganiczne). Atraktantem dla tlenowych i repelentem dla beztlenowych prokariontów jest tlen cząsteczkowy. Atraktanty są często reprezentowane przez substraty pokarmowe, chociaż nie wszystkie niezbędne dla organizmu substancje działają jak atraktanty. Ponadto nie wszystkie trujące substancje działają jako repelenty i nie wszystkie repelenty są szkodliwe. Tak więc bakterie nie są w stanie reagować na żadne związki, a jedynie na pewne i różne dla różnych bakterii.
W strukturach powierzchniowych komórki bakteryjnej znajdują się specjalne cząsteczki białka - receptory, które specyficznie łączą się z określonym chemoefektorem, podczas gdy cząsteczka chemoefektora nie zmienia się, ale zachodzą zmiany konformacyjne w cząsteczce receptora. Receptory rozmieszczone są nierównomiernie na całej powierzchni komórki i są skoncentrowane na jednym z biegunów. Stan receptora odzwierciedla pozakomórkowe stężenie odpowiedniego efektora.
Chemotaksja ma wartość adaptacyjną. Na przykład formy vibrio cholery z upośledzoną chemotaksją są mniej zjadliwe.
Aerotaksja- Bakterie potrzebujące tlenu cząsteczkowego gromadzą się wokół pęcherzyków powietrza, które wpadły pod szkiełko nakrywkowe.
Fototaksja- ruch w kierunku lub od światła, charakterystyczny dla bakterii fototroficznych, które wykorzystują światło jako źródło energii.
magnetotaksja- zdolność bakterii wodnych zawierających kryształy minerałów zawierających żelazo do pływania wzdłuż linii ziemskiego pola magnetycznego.
termotaksja- ruch w kierunku zmiany temperatury, który ma bardzo ważne dla niektórych bakterii chorobotwórczych.
Lepkość- zdolność reagowania na zmiany lepkości roztworu. Bakterie zwykle preferują pożywkę o większej lepkości, co ma duże znaczenie dla gatunków chorobotwórczych.
Ślizgające się bakterie. Zdolność do ślizgania się z małą prędkością (2–11 µm / s) po stałym lub lepkim podłożu stwierdzono u niektórych prokariotów, na przykład mykoplazmy.
Istnieje kilka hipotez wyjaśniających ruch ślizgowy. Według hipoteza napędu odrzutowego jest to spowodowane wydzielaniem śluzu przez liczne pory śluzowe w SC, w wyniku czego komórka jest odpychana od podłoża w kierunku przeciwnym do kierunku wydzielania śluzu. Według hipoteza fali biegnącej Ruch ślizgowy w ruchliwych formach wolnych od wiciowców jest związany z obecnością pomiędzy warstwą peptydoglikanu a zewnętrzną błoną CL cienkiej warstwy białkowej uporządkowanych włókienek, podobnej do włókien wici. „Uruchomiony” przez te struktury ruch obrotowy fibryli prowadzi do pojawienia się „wędrującej fali” (poruszających się mikroskopijnych wybrzuszeń CL) na powierzchni komórki, w wyniku czego komórka jest odpychana od podłoża. Wreszcie, u niektórych ślizgających się bakterii opisano struktury przypominające podstawowe ciała form wiciowych.
Funkcje wici:
1. Zapewnij przyczepność - etap początkowy proces zakaźny.
2. Zapewnij ruchliwość bakterii.
3. Określ specyficzność antygenową, jest to antygen H.
Wykrywanie wici:
1. Mikroskopia fazowo-kontrastowa preparatów natywnych (krople „rozkruszone” i „wiszące”). Mikroskopowo określa się ruchliwość w komórkach z codziennej hodowli. W celu odróżnienia ruchliwości od biernych ruchów Browna, do kropli badanej kultury dodaje się kroplę 5% wodnego roztworu fenolu, w tym przypadku ruch czynny ustaje.
2. Mikroskopia ciemnego pola preparatów natywnych.
3. Mikroskopia świetlna preparatów barwionych barwnikami lub metalami. Ponieważ wici bardzo łatwo ulegają uszkodzeniu podczas przygotowywania preparatu, metody te są rzadko stosowane w codziennej praktyce.
Wici są barwione przy użyciu komórek hodowanych na skosie agarowym. Komórki znajdujące się w pobliżu wody kondensacyjnej wybiera się za pomocą ezy bakteryjnej i ostrożnie przenosi do sterylnej wody destylowanej o tej samej temperaturze, co temperatura inkubacji bakterii na skośnym agarze, a bakterii nie strząsa się z ezy, ale ostrożnie zanurza w wodzie. Probówkę z bakteriami pozostawia się w temperaturze pokojowej na 30 minut. Stosuje się chemicznie czyste (przemyte w mieszaninie chromu) szkło, na które nanosi się 2-3 krople zawiesiny. Zawiesinę rozprowadza się po powierzchni szkła, delikatnie ją przechylając. Wysuszyć lek na powietrzu.
Wici są bardzo cienkie, więc można je wykryć tylko za pomocą specjalnego przetwarzania. Początkowo za pomocą wytrawiania uzyskuje się spęcznienie i zwiększenie ich wielkości, a następnie preparat barwi się, dzięki czemu stają się widoczne pod mikroskopem świetlnym.
Częściej używany metoda srebrzenia wg Morozowa (ryc. 10):
- preparat utrwala się roztworem lodowatego kwasu octowego przez 1 minutę, przemywa wodą;
- nałożyć roztwór garbnika (opalanie, zagęszczenie wici) przez 1 minutę, przemyć wodą;
- preparat poddaje się ogrzewaniu z roztworem impregnującym azotanu srebra przez 1-2 minuty, przemywa wodą, suszy i bada pod mikroskopem.
Mikroskopowo widoczne są ciemnobrązowe komórki i jaśniejsze wici.

Ryż. 10. Identyfikacja wici przez srebrzenie
Ryż. jedenaście. Identyfikacja wici
za pomocą mikroskopii elektronowej
Ryc. 4. Mikroskopia elektronowa preparatów osadzonych z metalami ciężkimi (ryc. 11).
5. Pośrednio - zgodnie z charakterem wzrostu bakterii przy wysianiu w półpłynnym 0,3% agarze. Po inkubacji upraw w termostacie przez 1–2 dni obserwuje się charakter rozwoju bakterii:
- u bakterii nieruchliwych (np. saprophyticus) wzdłuż wstrzyknięcia występuje narośl - „gwóźdź”, a podłoże jest przezroczyste;
u bakterii ruchliwych (np. E coli) występuje narośl z boku iniekcji, w całej kolumnie agarowej – „jodełka”, oraz rozproszone zmętnienie podłoża.
Wszystkie bakterie dzielą się na ruchome i nieruchome. Organami ruchu u bakterii są wici. Składają się z flageliny, białka, które jest strukturalnie spokrewnione z białkami kurczliwymi typu miozyny.
Podstawa wici to ciało podstawowe, składające się z układu krążków (blefaroplast: 1 krążek – zewnętrzna strona ściany komórkowej, 2 krążek – wewnętrzna strona ściany komórkowej, 3 krążek – błona cytoplazmatyczna), „osadzone” w cytoplazmie błona i ściana komórkowa. Długość wici jest większa niż długość ciała samego drobnoustroju.
Według liczby wici i ich lokalizacji, ruchliwe mikroorganizmy dzielą się na:
1. Monotrichous, posiadający jedną wić na końcu ciała (najbardziej ruchliwą). Na przykład Vibrio cholerae.
2. Lophotrichous, posiadający wiązkę wici na jednym z biegunów komórki. Na przykład Burkholderia (Pseudomonas) pseudomalei jest czynnikiem sprawczym melioidozy.
3. Amphitrichous, posiadający wici na obu biegunach komórki. Na przykład Spirillum volutans.
4. Peritrichous, posiadający wici na całym obwodzie komórki. Na przykład Escherichia coli, Salmonella typhi.
Wykrywanie wici. Wici są bardzo cienkie, więc można je wykryć tylko za pomocą specjalnego przetwarzania. W szczególności najpierw za pomocą zaprawy uzyskuje się pęcznienie i zwiększenie ich wielkości, a następnie preparat jest barwiony, dzięki czemu stają się widoczne pod mikroskopem świetlnym. Wici można wykryć za pomocą barwienia według Morozowa, Lefflera, a także za pomocą mikroskopii elektronowej. Wici można również wykryć na podstawie aktywnej ruchliwości bakterii.
Ruch drobnoustrojów obserwuje się w preparatach „rozgniecionych” i „wiszących” kropli z żywych kultur. Preparaty te są mikroskopowane suchą lub zanurzeniową soczewką w ciemnym polu lub w kontraście fazowym. Ponadto ruchliwość można określić na podstawie charakteru wzrostu bakterii w półpłynnym agarze.
Wypił z bakterii.
Pili (pili), synonimy: kosmki, fimbria, to cienkie puste w środku włókna o charakterze białkowym, które pokrywają powierzchnię komórek bakteryjnych. W przeciwieństwie do wici nie pełnią funkcji motorycznej.
Pili wystają z powierzchni komórki i składają się z białka pilina.
Zgodnie z ich przeznaczeniem funkcjonalnym dzielą się na 2 typy.
1) Pilusy pierwszego typu są obecne u większości bakterii, dlatego nazywane są „kosmkami typu ogólnego” (pilusy pospolite). Powodują przyczepianie się lub adhezję bakterii do określonych komórek organizmu gospodarza. Adhezja jest początkowym etapem każdego procesu zakaźnego.
2) Pilusy drugiego typu (synonimy: koniugacyjny lub seksualny - pilusy płciowe) są dostępne tylko u bakterii dawcy, które mają specjalny plazmid. Ich liczba jest niewielka - 1-4 na komórkę.
Piły podłogowe spełniają następujące funkcje:
1. Uczestniczyć w przenoszeniu materiału genetycznego z jednej komórki do drugiej podczas koniugacji bakterii.
2. Adsorbują określone wirusy bakteryjne – bakteriofagi
Spory bakteryjne, warunki powstawania, umiejscowienie, mechanizm i etapy barwienia Aujeszky'ego.
spór- szczególna forma bakterii spoczynkowych o Gram-dodatnim typie budowy ściany komórkowej.
sporulacja- jest to sposób zachowania gatunku (genofor) w środowisku zewnętrznym w niesprzyjających warunkach, a nie metoda rozmnażania.
Tworzą się zarodniki w niesprzyjających warunkach dla bytowania bakterii (przesuszenie, niedobór składników pokarmowych itp.). Wewnątrz komórki bakteryjnej powstaje jeden zarodnik (endospora).
Etapy powstawania zarodników
1. Przygotowawczy. W cytoplazmie bakterii tworzy się zagęszczony obszar, który nie ma wolnej wody, zwany „strefą zarodnikową”, który zawiera nukleoid.
2. Stadium prespor (prospor). Wokół strefy sporogennej tworzy się otoczka podwójnej błony cytoplazmatycznej.
3. Tworzenie kory składającej się z peptydoglikanu i błony zewnętrznej o wysokiej zawartości soli wapnia i lipidów.
4. Etap dojrzewania. Po zewnętrznej stronie błony zewnętrznej tworzy się otoczka zarodników, po czym wegetatywna część komórki ulega lizie, uwalniając zarodnik.
Rozwój mikrobiologii przyniósł w ostatnich dziesięcioleciach wiele odkryć. A jedną z nich jest specyfika ruchu wiciowców. Konstrukcja silników tych starożytnych organizmów okazała się bardzo złożona i zgodnie z zasadą ich pracy bardzo różni się od wici naszych najbliższych eukariotycznych krewnych pierwotniaków. Silnik wiciowców był najgorętszą kontrowersją między kreacjonistami a ewolucjonistami. O bakteriach, ich motorach wiciowych i nie tylko - ten artykuł.
Biologia ogólna
Na początek przypomnijmy sobie, jakie to organizmy i jakie miejsce zajmują w systemie organicznego świata naszej planety. Domena Bacteria zrzesza ogromną liczbę jednokomórkowych organizmów prokariotycznych (bez uformowanego jądra).
Te żywe komórki pojawiły się na scenie życia prawie 4 miliardy lat temu i były pierwszymi osadnikami planety. Mogą mieć bardzo różne kształty (ziarniaki, pręciki, wibratory, krętki), ale większość z nich jest wiciowata.
Gdzie żyją bakterie? Wszędzie. Na planecie żyje ponad 5 × 10 30. Jest ich około 40 milionów w 1 gramie gleby, aż 39 bilionów żyje w naszym organizmie. Można je znaleźć na dnie Rowu Mariańskiego, w gorących „czarnych palaczach” na dnie oceanów, w lodach Antarktydy, a na dłoniach masz obecnie nawet 10 milionów bakterii.
Wartość jest niezaprzeczalna
Pomimo ich mikroskopijnych rozmiarów (0,5-5 mikronów), ich całkowita biomasa na Ziemi jest większa niż biomasa zwierząt i roślin razem wziętych. Ich rola w obiegu substancji jest niezastąpiona, a ich właściwości konsumpcyjne (niszczyciele materii organicznej) nie pozwalają na zasypanie planety górami trupów.
Cóż, nie zapomnij o patogenach: czynnikami sprawczymi dżumy, ospy, kiły, gruźlicy i wielu innych chorób zakaźnych są również bakterie.
Bakterie znalazły zastosowanie w działalności gospodarczej człowieka. Począwszy od przemysłu spożywczego ( nabiał, sery, kiszone warzywa, napoje alkoholowe), „zielonej” gospodarki (biopaliwa i biogaz), po metody inżynierii komórkowej i produkcję leków (szczepionki, surowice, hormony, witaminy).

Morfologia ogólna
Jak już wspomniano, ci jednokomórkowi przedstawiciele życia nie mają jądra, ich materiał dziedziczny (cząsteczki DNA w postaci pierścienia) znajdują się w pewnym obszarze cytoplazmy (nukleoid). Ich komórka ma błonę plazmatyczną i gęstą kapsułkę utworzoną przez mureinę peptydoglikanu. Z organelli komórkowych bakterie mają mitochondria, mogą występować chloroplasty i inne struktury o różnych funkcjach.
Większość bakterii to wici. Ciasna kapsułka na powierzchni komórki zapobiega ich przemieszczaniu się poprzez zmianę samej komórki, tak jak robią to ameby. Ich wici to gęste formacje białkowe o różnej długości i średnicy około 20 nm. Niektóre bakterie mają pojedynczą wici (monotrych), podczas gdy inne mają dwie (amphitrichous). Czasami wici są ułożone w pęczki (lophotrichous) lub pokrywają całą powierzchnię komórki (peritrichous).
Wiele z nich żyje jako pojedyncze komórki, ale niektóre tworzą skupiska (pary, łańcuchy, włókna, strzępki).

Funkcje ruchu
Biczowane bakterie mogą poruszać się na różne sposoby. Niektóre poruszają się tylko do przodu i zmieniają kierunek, przewracając się. Niektóre są zdolne do drgań, inne poruszają się ślizgając.
Wici bakterii pełnią funkcje nie tylko komórkowego „wiosła”, ale mogą być również narzędziem „deskacyjnym”.
Do niedawna uważano, że wić bakterii macha jak ogon węża. Ostatnie badania wykazały, że wić bakterii jest znacznie bardziej skomplikowana. Działa jak turbina. Zamocowany do napędu obraca się w jednym kierunku. Siłownik lub silnik wiciowy bakterii to złożona struktura molekularna, która działa jak mięsień. Z tą różnicą, że mięsień po skurczu musi się rozluźnić, a motor bakteryjny pracuje cały czas.

Nanomechanizm wici
Nie zagłębiając się w biochemię ruchu, zauważamy, że w tworzeniu napędu wici bierze udział aż 240 białek, które podzielone są na 50 składników molekularnych o określonej funkcji w ustroju.
W tym systemie napędowym bakterii znajduje się wirnik, który się porusza i stojan, który zapewnia ten ruch. Jest wał napędowy, tuleja, sprzęgło, hamulce i akceleratory
Ten miniaturowy silnik pozwala bakteriom przemieszczać się 35 razy w stosunku do ich własnego rozmiaru w ciągu zaledwie 1 sekundy. Jednocześnie praca samej wici, która wykonuje 60 tysięcy obrotów na minutę, organizm zużywa tylko 0,1% całej energii, którą zużywa komórka.
Zaskakujące jest również to, że bakteria może wymieniać i naprawiać wszystkie części zamienne swojego mechanizmu silnikowego „w biegu”. Wyobraź sobie, że jesteś w samolocie. A technicy wymieniają łopatki pracującego silnika.
Bakteria wiciowata kontra Darwin
Silnik zdolny do pracy z prędkością do 60 000 obrotów na minutę, samoczynnie uruchamiający się i wykorzystujący jako paliwo wyłącznie węglowodany (cukier), posiadający urządzenie zbliżone do silnika elektrycznego - czy takie urządzenie mogło powstać w procesie ewolucji?
To pytanie zadał sobie dr biologii Michael Behe w 1988 roku. Wprowadził do biologii pojęcie systemu nieredukowalnego – systemu, w którym wszystkie jego części są jednocześnie niezbędne do zapewnienia jego działania, a usunięcie przynajmniej jednej części prowadzi do całkowitego zakłócenia jego funkcjonowania.
Z punktu widzenia ewolucji Darwina wszystkie zmiany strukturalne w ciele następują stopniowo i tylko te, które się powiodły, są wybierane przez dobór naturalny.
Wnioski M. Behe, przedstawione w książce „Czarna skrzynka Darwina” (1996): silnik bakterii wiciowej jest niepodzielnym systemem złożonym z ponad 40 części, a brak przynajmniej jednej doprowadzi do całkowitej dysfunkcji system, co oznacza, że system ten nie mógł powstać w wyniku doboru naturalnego.

Balsam dla kreacjonistów
Teoria stworzenia przedstawiona przez naukowca i profesora biologii, dziekana Wydziału Nauk Biologicznych na Lehigh University of Bethlehem (USA) M. Behe natychmiast przyciągnęła uwagę duchownych kościelnych i zwolenników teorii boskości pochodzenie życia.
W 2005 roku w Stanach Zjednoczonych doszło nawet do procesu, w którym Behe był świadkiem ze strony zwolenników teorii „rozsądnego projektu”, który rozważał wprowadzenie nauki kreacjonizmu w szkołach Dover w ramach kursu „ O pandach i ludziach”. Proces przegrał, nauczanie takiego przedmiotu uznano za sprzeczne z obowiązującą konstytucją.
Ale debata między kreacjonistami a ewolucjonistami trwa do dziś.
Wić to struktura powierzchniowa komórki bakteryjnej, która służy im do poruszania się w płynnych mediach.
W zależności od umiejscowienia wici bakterie dzielą się na (ryc. 1):
perytrychalny
mieszany
Polak
subpolarny
wici bieguna- jedna lub więcej wici znajduje się na jednym (monopolarnym) lub obu (dwubiegunowym) biegunie komórki, a podstawa jest równoległa do długiej osi komórki.
Wici subpolarne(subpolarny) - jedna lub więcej wici znajduje się w punkcie przejścia powierzchni bocznej do bieguna komórki na jednym lub dwóch jej końcach. U podstawy jest kąt prosty z dłuższą osią komórki.
Wici boczne(boczna) - jedna lub więcej wici w postaci wiązki znajduje się w środku jednej z połówek komórki.
Wici perytrychalne- rozmieszczone na całej powierzchni komórki pojedynczo lub w wiązkach, bieguny są zwykle ich pozbawione.
Mieszana wici- znajdują się dwie lub więcej wici różne punkty komórki.
W zależności od liczby wici wyróżnia się:
Monotrichous - jedna wici
Polytrichs - wiązka wici
Wyróżniono także:
lofotryczny- monoply politrychowy układ wici.
amfitryki- dwubiegunowy politrychowy układ wici.
Budowa wici bakteryjnej i ciała podstawowego. Rozłóg.
Sama wici jest ułożona dość prosto: włókno, które jest przymocowane do ciała podstawowego. Czasami pomiędzy korpus podstawowy a włókno można włożyć zakrzywiony odcinek rurki, tzw.
Pod względem składu chemicznego wić składa się w 98% z białka wici (flagellum - wici), zawiera 16 aminokwasów, przeważa glutamina i kwasy asparaginowy, tryptofan, cysteina i cystyna są nieobecne w niewielkiej ilości aminokwasów aromatycznych. Flagelina narzuca specyficzność antygenową, nazywana jest antygenem H. Wici bakteryjne nie mają aktywności ATPazy.
Grubość wici wynosi 10–12 nm, długość 3–15 µm.
Jest to sztywna spirala, skręcona w kierunku przeciwnym do ruchu wskazówek zegara. Obrót wici odbywa się również w kierunku przeciwnym do ruchu wskazówek zegara z częstotliwością od 40 obr./min do 60 obr./min, co powoduje, że komórka obraca się w przeciwnym kierunku, ale ponieważ komórka jest znacznie cięższa niż wici, wtedy jej rotacja jest wolniejsza od 12 do 14 obrotów na minutę.
Wić wyrasta z dystalnego końca, gdzie podjednostki wchodzą przez kanał wewnętrzny. U niektórych gatunków wić jest dodatkowo pokryta od zewnątrz osłonką, która jest kontynuacją ściany komórkowej i prawdopodobnie ma taką samą budowę.
Podstawowy korpus
Ciało podstawowe składa się z 4 części:
Łączenie pręta z żarnikiem lub haczykiem
Dwa dyski nawleczone na pręt. (M i S)
Grupa kompleksów białkowych (statorów)
czapeczka białkowa
Bakterie posiadające błonę wewnętrzną i zewnętrzną posiadają 2 dodatkowe krążki (P i L) oraz struktury białkowe, które znajdują się na błonie zewnętrznej w pobliżu podstawy ciała, stąd nie odgrywają istotnej roli w ruchu.
Specyfika struktury ciała podstawowego jest określona przez strukturę ściany komórkowej: jej nienaruszenie jest niezbędne do ruchu wici. Traktowanie komórek lizozymem prowadzi do usunięcia warstwy peptydoglikanu ze ściany komórkowej, co prowadzi do utraty ruchu, mimo że struktura wici nie została naruszona.
Wici bakteryjne determinują ruchliwość komórki bakteryjnej. Wici to cienkie włókna, które wywodzą się z błony cytoplazmatycznej i są dłuższe niż sama komórka. Wici mają grubość 12–20 nm i długość 3–15 µm. Składają się z 3 części: spiralnej nici, haczyka i korpusu podstawowego zawierającego pręcik ze specjalnymi krążkami (1 para krążków dla bakterii Gram-dodatnich i 2 pary krążków dla bakterii Gram-ujemnych). Krążki wici są przyczepione do błony cytoplazmatycznej i ściany komórkowej. Stwarza to efekt silnika elektrycznego z prętem silnikowym, który obraca wici. Wici składają się z białka - flageliny (od wici - wici); jest antygenem H. Podjednostki flageliny są zwinięte. Liczba wici w bakteriach różnych gatunków waha się od jednej (monotrich) u Vibrio cholerae do dziesięciu lub setek wici rozciągających się wzdłuż obwodu bakterii (peritrich) u Escherichia coli, Proteus itp. Lofotrich ma wiązkę wici w jednej koniec komórki. Amphitrichous mają jedną wici lub wiązkę wici na przeciwległych końcach komórki.
Pili (fimbrie, kosmki) - formacje nitkowate, cieńsze i krótsze (3-10 nm x 0,3-10 mikronów) niż wici. Pili wystają z powierzchni komórki i składają się z białka pilinowego, które ma aktywność antygenową. Istnieją pilusy odpowiedzialne za adhezję, czyli przyczepianie się bakterii do zaatakowanej komórki, a także pilusy odpowiedzialne za odżywianie, gospodarkę wodno-solną i seksualną (F-pilusy), czyli koniugacyjne. Napoje są obfite - kilkaset na klatkę. Jednak pilusy płciowe są zwykle 1-3 na komórkę: są one tworzone przez tak zwane „męskie” komórki dawcy zawierające przenośne plazmidy (plazmidy F-, R-, Col). Charakterystyczną cechą pilusów płciowych jest interakcja ze specjalnymi „męskimi” kulistymi bakteriofagami, które są intensywnie adsorbowane na pilusach płciowych.
Zarodniki to specyficzna forma uśpionych bakterii Firmicute, tj. bakterie o gram-dodatniej strukturze ściany komórkowej. Zarodniki powstają w warunkach niesprzyjających dla istnienia bakterii (suszenie, niedobór składników odżywczych itp.. Wewnątrz komórki bakteryjnej powstaje jeden zarodnik (endospora). Tworzenie zarodników przyczynia się do zachowania gatunku i nie jest metodą rozmnażania , podobnie jak u grzybów Bakterie tworzące przetrwalniki z rodzaju Bacillus mają zarodniki, które nie przekraczają średnicy komórki. Clostridium - wrzecionowate). Zarodniki są kwasoodporne, dlatego wybarwiają się na czerwono wg metody Aujeszky'ego lub wg metody Ziehla-Neelsena, a komórka wegetatywna na niebiesko.
Kształt sporu może być owalny, kulisty; lokalizacja w komórce jest terminalna, tj. na końcu sztyftu (w czynniku wywołującym tężec), subterminalny - bliżej końca sztyftu (w patogenach zatrucia jadem kiełbasianym, zgorzeli gazowej) i centralny (w prątkach wąglika). Zarodnik utrzymuje się przez długi czas ze względu na obecność wielowarstwowej otoczki, dipikolinianu wapnia, niskiej zawartości wody i powolnych procesów metabolicznych. W sprzyjających warunkach zarodniki kiełkują przez trzy kolejne etapy: aktywację, inicjację, kiełkowanie.
8. Główne formy bakterii
Bakterie kuliste (ziarniaki) są zwykle kuliste, ale mogą być lekko owalne lub w kształcie fasoli. Cocci można zlokalizować pojedynczo (mikrokoki); w parach (diplokoki); w postaci łańcuszków (paciorkowce) lub kiści winogron (gronkowce), pakiet (sarcinas). Paciorkowce mogą powodować zapalenie migdałków i różycę, gronkowce - różne procesy zapalne i ropne.
bakterie w kształcie pałeczek Najpopularniejszy. Pręciki mogą być pojedyncze, połączone w pary (diplobakterie) lub w łańcuchy (streptobakterie). Bakterie w kształcie pałeczek obejmują Escherichia coli, patogeny salmonellozy, czerwonki, duru brzusznego, gruźlicy itp. Niektóre bakterie w kształcie pałeczek mają zdolność tworzenia się w niesprzyjających warunkach sprzeczanie się. Pręciki tworzące zarodniki to tzw pałeczki. Nazywa się pałeczki wrzecionowate Clostridia.
Zarodnikowanie to złożony proces. Zarodniki znacznie różnią się od normalnej komórki bakteryjnej. Mają gęstą skorupę i bardzo małą ilość wody, nie wymagają składników odżywczych, a rozmnażanie całkowicie ustaje. Zarodniki są w stanie wytrzymać wysychanie, wysokie i niskie temperatury przez długi czas i mogą być zdolne do życia przez dziesiątki i setki lat (zarodniki wąglika, zatrucia jadem kiełbasianym, tężca itp.). W sprzyjającym środowisku zarodniki kiełkują, to znaczy zamieniają się w zwykłą wegetatywną formę rozmnażania.
Splecione bakterie może mieć postać przecinka - vibrios, z kilkoma lokami - spirilla, w postaci cienkiego skręconego sztyftu - krętków. Wibratory są czynnikiem sprawczym cholery, a czynnikiem sprawczym kiły jest krętek.
9. Cechy morfologii riketsji i chlamydii
Rickettsia to małe Gram-ujemne mikroorganizmy charakteryzujące się wyraźnym polimorfizmem - tworzą ziarniaki, formy w kształcie pręcików i nitkowate (ryc. 22). Rozmiary riketsji wahają się od 0,5 do 3-4 mikronów, długość form nitkowatych sięga 10-40 mikronów. Nie tworzą zarodników i kapsułek, według Zdrodowskiego są zabarwione na czerwono.
Chlamydia są kuliste, jajowate lub w kształcie pręta. Ich rozmiary wahają się w granicach 0,2-1,5 mikrona. Morfologia i wielkość chlamydii zależy od etapu ich wewnątrzkomórkowego cyklu rozwojowego, który charakteryzuje się przekształceniem małej kulistej formacji elementarnej w duże ciało początkowe z podziałem binarnym. Przed podziałem cząsteczki chlamydii otoczone są formacją przypominającą torebkę bakteryjną. Barwienie chlamydią "* według Romanovsky'ego-Giemsy, Gram-ujemne, wyraźnie widoczne w preparatach dożylnych z mikroskopem z kontrastem fazowym.
10. Budowa i biologia mykoplazm.
Tylko jeden rząd, Mycoplasmatales, należy do klasy Mollicutes. Przedstawiciele tego rzędu - mykoplazmy -
Różnią się od bakterii tym, że nie mają ściany komórkowej. Zamiast tego zawierają trójwarstwową błonę cytoplazmatyczną lipoprotein. Rozmiary mykoplazm wahają się w granicach 125-250 mikronów. Są okrągłe, owalne lub nitkowate, Gram-ujemne.
Mykoplazmy rozmnażają się przez rozszczepienie binarne, podobnie jak większość bakterii, zwłaszcza po utworzeniu małych formacji kokosowych (ciał elementarnych, EB) w strukturach włóknistych.
Mykoplazmy są zdolne do pączkowania i segmentacji. Minimalną jednostką odtwarzającą jest ET (0,7-0,2 mikrona). Głównym składnikiem błony komórkowej jest cholesterol. Mykoplazmy nie są zdolne do tworzenia cholesterolu i wykorzystują go z tkanek lub pożywek uzupełnionych ich wprowadzeniem. Barwienie Grama jest ujemne, ale barwienie Romanovsky'ego-Giemsy daje najlepsze wyniki. Mykoplazmy wymagają warunków hodowli: do pożywek należy dodać natywną surowicę, cholesterol, kwasy nukleinowe, węglowodany, witaminy i różne sole. Na gęstym podłożu tworzą charakterystyczne małe, przezroczyste kolonie z wypukłym, ziarnistym środkiem, nadając im wygląd „jajek sadzonych”. Na podłożach z krwią niektóre rodzaje mykoplazm powodują a- i beta-hemolizę. W półpłynnych pożywkach mykoplazmy rosną wzdłuż linii iniekcji, tworząc rozproszone, kruche kolonie. W mediach płynnych powodują lekkie zmętnienie lub opalescencję; niektóre szczepy są w stanie tworzyć najcieńszy tłusty film. U ludzi izolowano przedstawicieli rodzajów Mycoplasma, Ureaplasma i Acholeplasma, w tym gatunki chorobotwórcze i saprofityczne.