Selon l'emplacement des flagelles, les bactéries sont divisées en. Organelles du mouvement: fonctions et structure, caractéristiques du mouvement des protozoaires
La structure. Environ la moitié des espèces de bactéries connues à la surface ont des organes de mouvement - des flagelles incurvés en forme de vagues. La masse de flagelles représente jusqu'à 2% de la masse sèche de la bactérie. La longueur du flagelle est plus longue que la longueur du corps du micro-organisme et est de 3 à 12 μm; l'épaisseur du flagelle est de 0,02 μm et les flagelles polaires sont plus épais que les péritriches.
Les flagelles sont composées de protéine flagelline (flagelle latin - flagelle), qui dans sa structure appartient aux protéines contractiles de type myosine. Le flagelle contient soit un filament protéique homogène, soit 2 à 3 filaments étroitement pliés en une tresse. Le fil du flagelle est une spirale rigide torsadée dans le sens inverse des aiguilles d'une montre ; le pas de l'hélice est propre à chaque espèce bactérienne.
Le nombre, la taille et la disposition des flagelles sont des caractéristiques constantes pour une espèce particulière et sont prises en compte dans la taxonomie. Cependant, certaines bactéries peuvent former des flagelles. différents types. De plus, la présence de flagelles dépend des conditions environnementales : sur des milieux solides pendant une culture à long terme, les bactéries peuvent perdre des flagelles, et sur des milieux liquides, elles peuvent être acquises à nouveau. Le nombre et l'emplacement des flagelles dans la même espèce peuvent être déterminés par le stade du cycle de vie. Par conséquent, l'importance taxonomique de cette caractéristique ne doit pas être surestimée.
Classification des bactéries selon le nombre et la localisation des flagelles :
1. Atrichi - les flagelles sont absents.
2. Monotrichus- un flagelle situé à l'un des pôles de la cellule (genre Vibrio)- disposition monopolaire monotriche des flagelles, les bactéries les plus mobiles.
3. Polytrichus - nombreux flagelles :
– lophotriche- faisceau de flagelles à un pôle de la cellule (naissance Pseudomonas, Burkholderia) - arrangement polytriche monopolaire des flagelles;
– amphitriche- à chaque pôle de la cellule il y a un faisceau de flagelle (genre Spirillum)- disposition polytriche bipolaire des flagelles ;
– péritricheux- les flagelles sont disposés sans ordre particulier sur toute la surface de la cellule (fam. Entérobactéries(naissance Escherichia, Protée), fam. bacillacées, famille Clostidiacées), le nombre de flagelles varie de 6 à 1000 par cellule, selon le type de bactérie (Fig. 7).
Fig.7. Options pour la localisation des flagelles dans les bactéries :
1 - monotrich, 2 - lophotrich;
3 - amphitriches; 4 - péritrich.
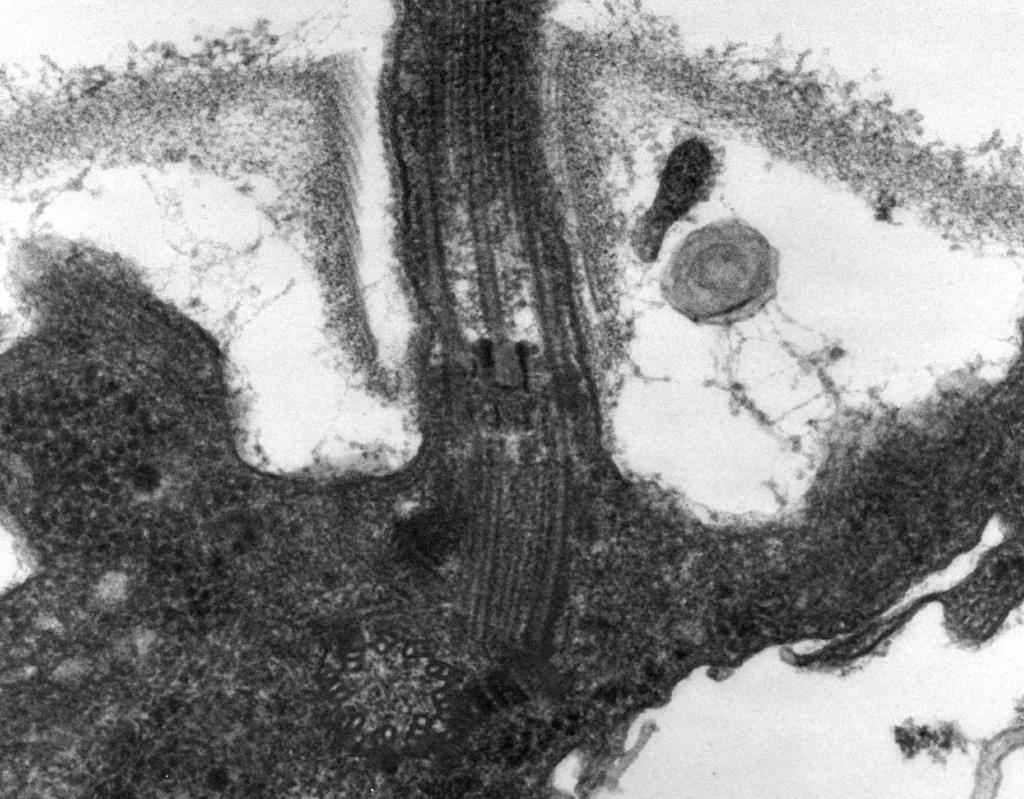
– La microscopie électronique révèle que le flagelle est composé de trois parties : filament en spirale, crochet et corps basal (Fig. 8).
La partie principale du flagelle est longue fil en spirale (fibrille) - un cylindre creux rigide d'un diamètre d'environ 120 nm, constitué de protéine flagelline. Les molécules de protéines forment 11 rangées sur la longueur du fil et sont disposées en spirale. Lors de la croissance du fil, les molécules de protéines synthétisées à l'intérieur de la cellule traversent la cavité du cylindre et sont attachées à une spirale à son extrémité. À l'extrémité du flagelle se trouve un capuchon protéique (couvercle) qui ferme l'ouverture du cylindre et empêche la libération de molécules protéiques dans l'environnement. La longueur du filament du flagelle peut atteindre plusieurs micromètres. Chez certaines espèces de bactéries, le flagelle est en outre recouvert d'une gaine à l'extérieur. À la surface du CS, le fil en spirale passe dans une structure incurvée épaissie - un crochet.

Riz. 8. Schéma de la structure du flagelle
2. Crochet(20–45 nm d'épaisseur) près de la surface cellulaire se trouve un cylindre relativement court, composé d'une protéine différente de la flagelline, et sert à fournir une connexion flexible du filament au corps basal.
3. Corps basal situé à la base du flagelle et assure sa rotation. Le corps basal contient 9 à 12 protéines différentes et se compose de deux ou quatre disques (anneaux) enfilés sur une tige, qui est une continuation du crochet. Ces anneaux sont montés dans le CPM et le CS. Les deux anneaux internes (M et S) sont des composants obligatoires du corps basal. L'anneau M est localisé dans le CPM, l'anneau S est situé dans l'espace périplasmique des bactéries Gram-négatives ou dans le sac peptidoglycane des bactéries Gram-positives. Les deux anneaux externes (D et L) sont facultatifs pour le mouvement, car ils ne sont présents que chez les bactéries Gram-négatives et sont localisés, respectivement, dans la couche de peptidoglycane et dans la membrane externe du CL. Les anneaux S, D et L sont immobiles et servent à fixer le flagelle dans le CS. La rotation du flagelle est déterminée par la rotation de l'anneau M intégré au CPM de la cellule. Ainsi, les caractéristiques structurelles du corps basal du flagelle sont déterminées par la structure du CS.
Fonctionnellement, le corps basal est un moteur électrique alimenté par des protons. L'anneau M du corps basal (rotor rotatif) est entouré de protéines membranaires à charges négatives (stator moteur). La cellule bactérienne possède un mécanisme efficace pour convertir l'énergie électrochimique en énergie mécanique. Par conséquent, la bactérie dépense environ 0,1% de toute l'énergie qu'elle dépense pour le fonctionnement du flagelle. Lorsque le flagelle fonctionne, une force proton-motrice est utilisée, qui est fournie par la différence de concentrations de protons sur les côtés externe et interne de la membrane (il y en a plus sur le côté externe) et la présence d'une charge plus négative sur la face interne de la membrane. La force motrice des protons oblige les protons à traverser le corps basal dans la cellule, tandis qu'ils s'attardent dans certaines parties du rotor, leur donnant une charge positive, puis les protons pénètrent à l'intérieur de la cellule. Les sections chargées sont situées de telle manière qu'une force d'attraction apparaît entre les sections chargées du rotor et du stator, l'anneau M commence à tourner à une vitesse d'environ 300 tr/min. Mécanisme de rotation : charge-recharge du groupe COOH dans les acides aminés. Pour une révolution complète de l'anneau, 500 à 1000 protons doivent traverser le corps basal. La rotation de l'anneau en M à travers l'axe relié rigidement à celui-ci et au crochet est transmise au filament du flagelle, qui fonctionne comme une hélice ou une hélice de navire. La bactérie nage tant que la vis fonctionne, la contribution de l'inertie est extrêmement faible.
De plus, les bactéries, même mortes, du milieu aquatique se déplacent sous l'effet du mouvement brownien. La cellule bactérienne est constamment soumise aux chocs des molécules environnantes, qui sont en mouvement thermique. Des coups provenant de différentes directions projettent la bactérie d'un côté à l'autre.
Le type de mouvement des flagelles est rotatif. Il existe deux types de mouvement : rectiligne et tumbling (changements aléatoires périodiques de direction de mouvement). Lorsque les flagelles tournent dans le sens antihoraire (environ 1 seconde), à une fréquence de 40 à 60 tr/min (proche de la vitesse d'un moteur électrique moyen), leurs fils sont tissés en un seul faisceau (Fig. 9a). La rotation des flagelles est transmise à la cellule. Comme la cellule est beaucoup plus massive que le flagelle, elle commence à se déplacer en ligne droite dans la direction opposée, à une vitesse 3 fois inférieure à la vitesse du flagelle.
Ceci assure le mouvement de translation de la cellule dont la vitesse en milieu liquide pour différents types bactéries est de 20 à 200 μm/s (cela correspond à environ 300 à 3 000 longueurs corporelles par minute) et un mouvement plus lent à la surface des milieux solides.
La bactérie peut nager délibérément dans une direction pendant 3 s au maximum, puis les impacts des molécules environnantes la font tourner dans une direction aléatoire. Par conséquent, les bactéries ont développé un mécanisme pour un changement spontané de la direction du mouvement - la commutation du moteur flagellaire. Lorsqu'elle commence à tourner dans le sens des aiguilles d'une montre (environ 0,1 s), la bactérie s'arrête et bascule (effectue une « culbute ») dans une direction aléatoire. Dans ce cas, les flagelles se dispersent dans différentes directions (Fig. 9b). Chez les amphitriches, lors du déplacement, un faisceau de flagelles est retourné (comme un parapluie retourné par le vent). Ensuite, le moteur passe à nouveau en rotation dans le sens inverse des aiguilles d'une montre et la bactérie nage à nouveau en ligne droite, mais dans une direction différente et aléatoire.
Les flagelles peuvent également changer de direction en réponse à un stimulus externe. Si la bactérie se déplace vers la concentration optimale en attractif, les flagelles poussent la cellule à travers le milieu, son mouvement rectiligne devient plus long et la fréquence de culbutage est plus faible, ce qui lui permet finalement de se déplacer dans la direction souhaitée.
Des cas d'existence de flagelles inactifs (paralysés) sont connus. Pour le mouvement des bactéries flagellées, l'intégrité (intégrité) du CS est nécessaire. Le traitement des cellules avec du lysozyme, qui entraîne l'élimination de la couche de peptidoglycane du CS, entraîne la perte de la capacité de la bactérie à se déplacer, bien que les flagelles restent intactes.
Taxis de bactéries. Au revoir Environnement reste inchangé, les bactéries nagent au hasard. Cependant, l'environnement est rarement complètement homogène. Si l'environnement est hétérogène, les bactéries présentent des réactions comportementales élémentaires : elles se déplacent activement dans la direction déterminée par certains facteurs externes. Ces mouvements intentionnels de bactéries déterminés génétiquement sont appelés taxis. Selon le facteur, on distingue la chimiotaxie (un cas particulier est l'aérotaxie), la phototaxie, la magnétotaxie, la thermotaxie et la viscositaxis.
Chimiotaxie- mouvement dans une certaine direction par rapport à la source du produit chimique. Substances chimiques divisé en deux groupes: taxis inertes et causants - chimioeffecteurs. Parmi les chimioeffecteurs, il existe des substances qui attirent les bactéries - attractifs (sucres, acides aminés, vitamines, nucléotides) et des substances qui les repoussent - répulsifs (certains acides aminés, alcools, phénols, ions inorganiques). L'attractif pour les aérobies et le répulsif pour les procaryotes anaérobies est l'oxygène moléculaire. Les attractifs sont souvent représentés par des substrats alimentaires, bien que toutes les substances nécessaires à l'organisme n'agissent pas comme des attractifs. De plus, toutes les substances toxiques ne servent pas de répulsifs et tous les répulsifs ne sont pas nocifs. Ainsi, les bactéries ne sont capables de réagir à aucun composé, mais seulement à certains et différents pour différentes bactéries.
Dans les structures de surface d'une cellule bactérienne, il existe des molécules protéiques spéciales - des récepteurs qui se connectent spécifiquement à un certain chimioeffecteur, tandis que la molécule chimioeffectrice ne change pas, mais des changements conformationnels se produisent dans la molécule réceptrice. Les récepteurs sont répartis de manière inégale sur toute la surface de la cellule et sont concentrés à l'un des pôles. L'état du récepteur reflète la concentration extracellulaire de l'effecteur correspondant.
La chimiotaxie a une valeur adaptative. Par exemple, les formes de vibrion cholérique à chimiotaxie altérée sont moins virulentes.
Aérotaxie- Les bactéries qui ont besoin d'oxygène moléculaire s'accumulent autour des bulles d'air tombées sous une lamelle.
Phototaxis- mouvement vers ou loin de la lumière, caractéristique des bactéries phototrophes qui utilisent la lumière comme source d'énergie.
magnétotaxie- la capacité des bactéries aquatiques contenant des cristaux de minéraux contenant du fer à nager le long des lignes du champ magnétique terrestre.
Thermotaxis- mouvement dans le sens du changement de température, qui a grande importance pour certaines bactéries pathogènes.
Viscositaxie- la capacité à réagir aux changements de viscosité de la solution. Les bactéries tendent généralement vers un milieu à viscosité plus élevée, ce qui est d'une grande importance pour les espèces pathogènes.
Bactéries glissantes. La capacité de glisser à faible vitesse (2 à 11 µm/s) sur un substrat solide ou visqueux a été trouvée chez certains procaryotes, par exemple les mycoplasmes.
Plusieurs hypothèses expliquent le mouvement de glissement. Selon hypothèse de propulsion à réaction il est dû à la sécrétion de mucus à travers de nombreux pores de mucus dans le SC, à la suite de quoi la cellule est repoussée du substrat dans la direction opposée à la direction de la sécrétion de mucus. Selon l'hypothèse de l'onde progressive le mouvement de glissement dans les formes mobiles sans flagelles est associé à la présence entre la couche de peptidoglycane et la membrane externe du CL d'une fine couche protéique de fibrilles ordonnées, similaires aux filaments des flagelles. Le mouvement de rotation des fibrilles, "lancé" par ces structures, conduit à l'apparition d'une "onde progressive" (renflements microscopiques mobiles du CL) à la surface de la cellule, à la suite de quoi la cellule est repoussée du substrat. Enfin, des structures ressemblant aux corps basaux des formes flagellées ont été décrites chez certaines bactéries glissantes.
Fonctions des flagelles :
1. Fournir une adhérence - stade initial processus infectieux.
2. Fournir la motilité des bactéries.
3. Déterminer la spécificité antigénique, c'est l'antigène H.
Détection des flagelles :
1. Microscopie à contraste de phase de préparations natives (gouttes « écrasées » et « suspendues »). Au microscope, la mobilité est déterminée dans les cellules de la culture quotidienne. Afin de distinguer la mobilité du mouvement brownien passif, une goutte d'une solution aqueuse de phénol à 5 % est ajoutée à une goutte de la culture étudiée ; dans ce cas, le mouvement actif s'arrête.
2. Microscopie à fond noir de préparations natives.
3. Microscopie optique de préparations colorées avec des colorants ou des métaux. Les flagelles étant très facilement endommagés lors de la préparation de la préparation, ces méthodes sont rarement utilisées dans la pratique quotidienne.
Les flagelles sont colorés à l'aide de cellules cultivées sur une gélose inclinée. Les cellules situées près de l'eau de condensation sont sélectionnées avec une boucle bactérienne et soigneusement transférées dans de l'eau distillée stérile à la même température que la température d'incubation des bactéries sur gélose inclinée, et les bactéries ne sont pas secouées de la boucle, mais soigneusement immergées dans l'eau. Le tube contenant les bactéries est laissé à température ambiante pendant 30 minutes. Du verre chimiquement pur (lavé dans un mélange de chrome) est utilisé, sur lequel 2-3 gouttes de suspension sont appliquées. La suspension est répartie sur la surface du verre en l'inclinant doucement. Séchez le médicament à l'air libre.
Les flagelles sont très fins, ils ne peuvent donc être détectés qu'avec un traitement spécial. Initialement, à l'aide de la gravure, un gonflement et une augmentation de leur taille sont obtenus, puis la préparation est colorée, grâce à quoi ils deviennent visibles au microscope optique.
Plus souvent utilisé méthode d'argenture selon Morozov (Fig. 10):
- la préparation est fixée avec une solution d'acide acétique glacial pendant 1 minute, lavée à l'eau ;
- appliquer une solution de tanin (tanin, rendant les flagelles plus denses) pendant 1 minute, lavé à l'eau ;
- la préparation est traitée par chauffage avec une solution d'imprégnation de nitrate d'argent pendant 1 à 2 minutes, lavée à l'eau, séchée et microscopique.
Au microscope, des cellules brun foncé et des flagelles plus clairs sont visibles.

Riz. dix. Identification des flagelles par argenture
Riz. Onze. Identification des flagelles
par microscopie électronique
Fig. 4. Microscopie électronique de préparations déposées avec des métaux lourds (Fig. 11).
5. Indirectement - selon la nature de la croissance bactérienne lorsqu'il est semé dans de la gélose semi-liquide à 0,3%. Après incubation des cultures dans un thermostat pendant 1 à 2 jours, la nature de la croissance bactérienne est notée:
- dans les bactéries non mobiles (ex. S. saprophyticus) il y a une croissance le long de l'injection - un «clou» et le milieu est transparent;
dans les bactéries mobiles (par ex. E. coli) il y a une croissance du côté de l'injection, dans toute la colonne de gélose - "chevrons", et une turbidité diffuse du milieu.
Toutes les bactéries divisé en mobile et immobile. Les organes de mouvement des bactéries sont les flagelles. Ils sont constitués de flagelline, une protéine structurellement apparentée aux protéines contractiles de type myosine.
Base du flagelle est un corps basal, constitué d'un système de disques (blépharoplaste: 1 disque - la face externe de la paroi cellulaire, 2 disque - la face interne de la paroi cellulaire, 3 disque - la membrane cytoplasmique), "incrusté" dans le cytoplasme membrane et paroi cellulaire. La longueur du flagelle est supérieure à la longueur du corps du microbe lui-même.
Selon le nombre de flagelles et leur emplacement, les micro-organismes mobiles sont divisés en :
1. Monotriches, ayant un flagelle à l'extrémité du corps (le plus mobile). Par exemple, Vibrio cholerae.
2. Lophotriches, ayant un faisceau de flagelles à l'un des pôles de la cellule. Par exemple, Burkholderia (Pseudomonas) pseudomalei est l'agent causal de la mélioïdose.
3. Amphitriches, ayant un flagelle aux deux pôles de la cellule. Par exemple, Spirillum volutans.
4. Péritriches, ayant des flagelles sur tout le périmètre de la cellule. Par exemple, Escherichia coli, Salmonella typhi.
Détection des flagelles. Les flagelles sont très fins, ils ne peuvent donc être détectés qu'avec un traitement spécial. En particulier, dans un premier temps, à l'aide d'un mordant, un gonflement et une augmentation de leur taille sont obtenus, puis la préparation est colorée, grâce à quoi ils deviennent visibles au microscope optique. Les flagelles peuvent être détectés par coloration selon Morozov, Leffler, ainsi que par microscopie électronique. Les flagelles peuvent également être détectés par la motilité active des bactéries.
Le mouvement des microbes est observé dans les préparations de gouttes "écrasées" et "suspendues" provenant de cultures vivantes. Ces préparations sont microscopiques avec une lentille sèche ou à immersion en fond noir ou en contraste de phase. De plus, la motilité peut être déterminée par la nature de la croissance des bactéries dans l'agar semi-liquide.
A bu des bactéries.
Pili (pili), synonymes : villosités, fimbria, sont de minces filaments creux de nature protéique qui recouvrent la surface des cellules bactériennes. Contrairement aux flagelles, ils n'exercent pas de fonction motrice.
Les pili s'étendent de la surface cellulaire et sont composés de protéines pilina.
Selon leur objectif fonctionnel, ils sont divisés en 2 types.
1) Les pili du premier type sont présents dans la plupart des bactéries, ils sont donc appelés « villosités de type général » (pili commun). Ils provoquent la fixation ou l'adhésion des bactéries à certaines cellules de l'organisme hôte. L'adhésion est l'étape initiale de tout processus infectieux.
2) Les pili du deuxième type (synonymes : conjugatif ou sexuel - pili sexuel) ne sont disponibles que chez les bactéries donneuses qui ont un plasmide spécial. Leur nombre est petit - 1 à 4 par cellule.
Les scies à sol remplissent les fonctions suivantes :
1. Participer au transfert de matériel génétique d'une cellule à une autre lors de la conjugaison des bactéries.
2. Ils adsorbent des virus bactériens spécifiques - les bactériophages
Spores bactériennes, conditions de formation, localisation, mécanisme et étapes de la coloration d'Aujeszky.
controverse- une forme particulière de bactérie au repos avec une structure de paroi cellulaire de type gram-positif.
sporulation- il s'agit d'un moyen de préservation d'une espèce (génophore) dans le milieu extérieur dans des conditions défavorables, et non d'un mode de reproduction.
Les spores se forment dans des conditions défavorables à l'existence de bactéries (assèchement, carence en nutriments, etc.). À l'intérieur de la cellule bactérienne, une spore (endospore) est formée.
Étapes de la formation des spores
1. Préparatoire. Dans le cytoplasme des bactéries, il se forme une zone compactée dépourvue d'eau libre, appelée "zone sporogène", qui contient le nucléoïde.
2. Stade de préspores (prospores). Autour de la zone sporogène, une gaine d'une double membrane cytoplasmique se forme.
3. Formation d'un cortex constitué de peptidoglycane et d'une membrane externe à haute teneur en sels de calcium et en lipides.
4. Stade de maturation. Une enveloppe de spores se forme sur la face externe de la membrane externe, après quoi la partie végétative de la cellule est lysée, libérant la spore.
Le développement de la microbiologie a apporté de nombreuses découvertes au cours des dernières décennies. Et l'un d'eux est les particularités du mouvement des bactéries flagellées. La conception des moteurs de ces organismes anciens s'est avérée très complexe et, selon le principe de leur travail, est très différente des flagelles de nos plus proches parents eucaryotes des protozoaires. Le moteur de la bactérie flagellée a été la controverse la plus chaude entre les créationnistes et les évolutionnistes. À propos des bactéries, de leurs moteurs flagellaires et bien plus encore - cet article.
Biologie générale
Pour commencer, rappelons quel genre d'organismes ils sont et quelle place ils occupent dans le système du monde organique sur notre planète. Le domaine des bactéries réunit un grand nombre d'organismes procaryotes unicellulaires (sans noyau formé).
Ces cellules vivantes sont apparues sur la scène de la vie il y a près de 4 milliards d'années et ont été les premiers colons de la planète. Ils peuvent être de formes très différentes (cocci, bâtonnets, vibrions, spirochètes), mais la plupart d'entre eux sont flagellés.
Où vivent les bactéries ? Partout. Plus de 5 × 10 30 vivent sur la planète. Il y en a environ 40 millions dans 1 gramme de sol, jusqu'à 39 billions vivent dans notre corps. On les trouve au fond de la fosse des Mariannes, dans des « fumeurs noirs » chauds au fond des océans, dans la glace de l'Antarctique, et vous avez actuellement jusqu'à 10 millions de bactéries sur les mains.
La valeur est indéniable
Malgré leur taille microscopique (0,5-5 microns), leur biomasse totale sur Terre est supérieure à la biomasse des animaux et des plantes réunis. Leur rôle dans la circulation des substances est irremplaçable, et leurs propriétés de consommateurs (destructeurs de matière organique) ne permettent pas de recouvrir la planète de montagnes de cadavres.
Eh bien, n'oubliez pas les agents pathogènes: les agents responsables de la peste, de la variole, de la syphilis, de la tuberculose et de nombreuses autres maladies infectieuses sont également des bactéries.
Les bactéries ont trouvé une application dans l'activité économique humaine. A partir de l'industrie alimentaire ( les produits laitiers, fromages, légumes marinés, boissons alcoolisées), de l'économie « verte » (biocarburants et biogaz) aux méthodes d'ingénierie cellulaire et de production de médicaments (vaccins, sérums, hormones, vitamines).

Morphologie générale
Comme déjà mentionné, ces représentants unicellulaires de la vie n'ont pas de noyau, leur matériel héréditaire (molécules d'ADN sous forme d'anneau) est situé dans une certaine zone du cytoplasme (nucléoïde). Leur cellule possède une membrane plasmique et une capsule dense formée de peptidoglycane muréine. Parmi les organites cellulaires, les bactéries ont des mitochondries, il peut y avoir des chloroplastes et d'autres structures ayant des fonctions différentes.
La plupart des bactéries sont des flagelles. La capsule serrée à la surface de la cellule les empêche de se déplacer en modifiant la cellule elle-même, comme le font les amibes. Leurs flagelles sont des formations protéiques denses de différentes longueurs et d'environ 20 nm de diamètre. Certaines bactéries ont un seul flagelle (monotriches), tandis que d'autres en ont deux (amphitriches). Parfois, les flagelles sont disposés en faisceaux (lophotriches) ou recouvrent toute la surface de la cellule (péritriches).
Beaucoup d'entre eux vivent comme des cellules uniques, mais certains forment des grappes (paires, chaînes, filaments, hyphes).

Caractéristiques du mouvement
Les bactéries flagellées peuvent se déplacer de différentes manières. Certains ne font qu'avancer, et changent de direction en culbutant. Certains sont capables de se contracter, d'autres se déplacent en glissant.
Les flagelles des bactéries remplissent non seulement les fonctions d'une "rame" cellulaire, mais peuvent également être un outil "d'embarquement".
Jusqu'à récemment, on croyait que le flagelle d'une bactérie remuait comme la queue d'un serpent. Des études récentes ont montré que le flagelle des bactéries est beaucoup plus compliqué. Il fonctionne comme une turbine. Attaché au lecteur, il tourne dans un sens. L'actionneur, ou moteur flagellaire des bactéries, est une structure moléculaire complexe qui fonctionne comme un muscle. A la différence que le muscle doit se détendre après la contraction, et que le moteur bactérien fonctionne en permanence.

Le nanomécanisme du flagelle
Sans entrer dans la biochimie du mouvement, nous notons que jusqu'à 240 protéines sont impliquées dans la création de la pulsion du flagelle, qui sont divisées en 50 composants moléculaires ayant une fonction spécifique dans le système.
Dans ce système de propulsion des bactéries, il y a un rotor qui se déplace et un stator qui assure ce mouvement. Il y a un arbre de transmission, une douille, un embrayage, des freins et des accélérateurs
Ce moteur miniature permet aux bactéries de voyager 35 fois leur propre taille en seulement 1 seconde. Dans le même temps, le travail du flagelle lui-même, qui fait 60 000 tours par minute, le corps ne dépense que 0,1% de toute l'énergie consommée par la cellule.
Il est également surprenant que la bactérie puisse remplacer et réparer toutes les pièces détachées de son mécanisme moteur « à la volée ». Imaginez que vous êtes dans un avion. Et les techniciens changent les pales d'un moteur en marche.
Bactérie flagellée vs Darwin
Un moteur capable de fonctionner à des vitesses allant jusqu'à 60 000 tours par minute, auto-démarrant et n'utilisant que des glucides (sucre) comme carburant, ayant un dispositif semblable à un moteur électrique - un tel dispositif aurait-il pu apparaître au cours du processus d'évolution ?
C'est la question que Michael Behe, docteur en biologie, s'est posée en 1988. Il a introduit en biologie le concept de système irréductible - un système dans lequel toutes ses parties sont simultanément nécessaires pour assurer son fonctionnement, et la suppression d'au moins une partie conduit à une perturbation complète de son fonctionnement.
Du point de vue de l'évolution de Darwin, tous les changements structurels dans le corps se produisent progressivement et seuls ceux qui réussissent sont sélectionnés par la sélection naturelle.
Les conclusions de M. Behe, exposées dans le livre "Darwin's Black Box" (1996): le moteur d'une bactérie flagellaire est un système indivisible de plus de 40 parties, et l'absence d'au moins une entraînera un dysfonctionnement complet de la système, ce qui signifie que ce système n'a pas pu se produire par sélection naturelle.

Baume pour les créationnistes
La théorie de la création telle que présentée par le scientifique et professeur de biologie, doyen de la Faculté des sciences biologiques de l'Université Lehigh de Bethléem (États-Unis) M. Behe a immédiatement attiré l'attention des ministres de l'Église et des partisans de la théorie du divin origine de la vie.
En 2005, aux États-Unis, il y a même eu un procès où Behe a été témoin des partisans de la théorie du "design raisonnable", qui envisageait l'introduction de l'étude du créationnisme dans les écoles de Douvres dans le cours " Sur les pandas et les gens." Le processus était perdu, l'enseignement d'une telle matière était reconnu comme contraire à la constitution en vigueur.
Mais le débat entre créationnistes et évolutionnistes se poursuit aujourd'hui.
Le flagelle est la structure de surface d'une cellule bactérienne, qui leur sert à se déplacer dans les milieux liquides.
Selon l'emplacement des flagelles, les bactéries sont divisées en (Fig. 1):
péritriche
mixte
Pôle
Subpolaire
Flagelles polaires- un ou plusieurs flagelles sont situés sur un (monopolaire) ou les deux pôles (bipolaire) de la cellule et la base est parallèle au grand axe de la cellule.
Flagelles subpolaires(subpolaire) - un ou plusieurs flagelles sont situés au point de transition de la surface latérale au pôle de la cellule à une ou deux de ses extrémités. A la base se trouve un angle droit avec le grand axe de la cellule.
Flagelles latéraux(latéral) - un ou plusieurs flagelles en forme de faisceau sont situés au milieu de l'une des moitiés de la cellule.
Flagelles péritriches- situés sur toute la surface de la cellule un à un ou en faisceaux, les pôles en sont généralement dépourvus.
Flagelles mixtes- deux ou plusieurs flagelles sont situés dans points différents cellules.
Selon le nombre de flagelles, il y a :
Monotriches - un flagelle
Polytrichs - bouquet de flagelles
Distinguait également :
lophotriche- disposition polytrichiale monopolistique des flagelles.
amphitriches- arrangement polytrichial bipolaire des flagelles.
La structure du flagelle bactérien et du corps basal. Flagelle.
Le flagelle lui-même est disposé assez simplement : un filament qui est attaché au corps basal. Parfois, une section courbe du tube, appelée crochet, peut être insérée entre le corps basal et le filament ; elle est plus épaisse que le filament et participe à la fixation flexible du filament au corps basal.
En termes de composition chimique, le flagelle est constitué à 98% de protéine flagelline (flagelle - flagelle), il contient 16 acides aminés, la glutamine et les acides aspartiques prédominent, le tryptophane, la cystéine et la cystine sont absents dans une petite quantité d'acides aminés aromatiques. La flagelline impose une spécificité antigénique, on l'appelle l'antigène H. Les flagelles bactériens n'ont pas d'activité ATPase.
L'épaisseur du flagelle est de 10–12 nm, la longueur est de 3–15 µm.
C'est une spirale rigide, tordue dans le sens antihoraire. La rotation du flagelle s'effectue également dans le sens inverse des aiguilles d'une montre avec une fréquence de 40 tr/min à 60 tr/min, ce qui fait tourner la cellule dans le sens opposé, mais puisque la cellule est beaucoup plus lourde que le flagelle, alors sa rotation est plus lente de 12 à 14 tr/min.
Le flagelle se développe à partir de l'extrémité distale, où les sous-unités pénètrent par le canal interne. Chez certaines espèces, le flagelle est en outre recouvert à l'extérieur d'une gaine, qui est une continuation de la paroi cellulaire et a probablement la même structure.
Corps basal
Le corps basal est composé de 4 parties :
Accouplement de tige avec filament ou crochet
Deux disques enfilés sur une tige. (M et S)
Groupe de complexes protéiques (stators)
bouchon de protéines
Les bactéries qui ont une membrane interne et externe ont 2 disques supplémentaires (P et L) et des structures protéiques qui se trouvent sur la membrane externe près du corps basal, elles ne jouent donc pas un rôle important dans le mouvement.
La particularité de la structure du corps basal est déterminée par la structure de la paroi cellulaire: son intégrité est nécessaire au mouvement des flagelles. Le traitement des cellules avec du lysozyme entraîne l'élimination de la couche de peptidoglycane de la paroi cellulaire, ce qui entraîne une perte de mouvement, bien que la structure du flagelle n'ait pas été perturbée.
Les flagelles bactériens déterminent la motilité de la cellule bactérienne. Les flagelles sont de minces filaments qui proviennent de la membrane cytoplasmique et sont plus longs que la cellule elle-même. Les flagelles ont une épaisseur de 12 à 20 nm et une longueur de 3 à 15 µm. Ils se composent de 3 parties : un fil en spirale, un crochet et un corps basal contenant une tige avec des disques spéciaux (1 paire de disques pour les bactéries gram-positives et 2 paires de disques pour les bactéries gram-négatives). Les disques des flagelles sont attachés à la membrane cytoplasmique et à la paroi cellulaire. Cela crée l'effet d'un moteur électrique avec une tige motrice qui fait tourner le flagelle. Les flagelles consistent en une protéine - la flagelline (de flagelle - flagelle); est un antigène H. Les sous-unités flagellines sont enroulées. Le nombre de flagelles dans les bactéries de différentes espèces varie d'un (monotrich) chez Vibrio cholerae à dix ou centaines de flagelles s'étendant le long du périmètre de la bactérie (peritrich) chez Escherichia coli, Proteus, etc. Les Lofotrichs ont un faisceau de flagelles à un fin de la cellule. Les amphitriches ont un flagelle ou un faisceau de flagelles aux extrémités opposées de la cellule.
Pili (fimbriae, villosités) - formations filamenteuses, plus fines et plus courtes (3-10 nm x 0,3-10 microns) que les flagelles. Les pili s'étendent de la surface cellulaire et sont constitués de la protéine piline, qui a une activité antigénique. Il existe des pili responsables de l'adhésion, c'est-à-dire de la fixation des bactéries à la cellule affectée, ainsi que des pili responsables de la nutrition, du métabolisme eau-sel et sexuel (F-pili), ou pili de conjugaison. Les boissons sont abondantes - plusieurs centaines par cage. Cependant, les pili sexuels sont généralement au nombre de 1 à 3 par cellule : ils sont formés par des cellules donneuses dites "mâles" contenant des plasmides transmissibles (F-, R-, Col-plasmids). Une caractéristique distinctive des pili sexuels est l'interaction avec des bactériophages sphériques "mâles" spéciaux, qui sont intensément adsorbés sur les pili sexuels.
Les spores sont une forme particulière de bactéries firmicutes dormantes, c'est-à-dire bactéries à structure de paroi cellulaire gram-positive. Les spores se forment dans des conditions défavorables à l'existence des bactéries (assèchement, carence en nutriments, etc.. Une spore (endospore) se forme à l'intérieur de la cellule bactérienne. La formation de spores contribue à la préservation de l'espèce et n'est pas un mode de reproduction , comme dans les champignons.Les bactéries sporulées du genre Bacillus ont des spores qui ne dépassent pas le diamètre de la cellule.Les bactéries dont la taille des spores dépasse le diamètre de la cellule sont appelées clostridies, par exemple, les bactéries du genre Clostridium (lat. Clostridium - fuseau).Les spores sont résistantes aux acides, elles sont donc colorées en rouge selon la méthode Aujeszky ou selon la méthode Ziehl-Neelsen, et la cellule végétative en bleu.
La forme du différend peut être ovale, sphérique; l'emplacement dans la cellule est terminal, c'est-à-dire à l'extrémité du bâton (dans l'agent causal du tétanos), subterminal - plus près de l'extrémité du bâton (dans les agents pathogènes du botulisme, la gangrène gazeuse) et central (dans les bacilles de l'anthrax). La spore persiste longtemps en raison de la présence d'une coquille multicouche, de dipicolinate de calcium, d'une faible teneur en eau et de processus métaboliques lents. Dans des conditions favorables, les spores germent en trois étapes successives : activation, initiation, germination.
8. Les principales formes de bactéries
Bactéries globulaires (cocci) sont généralement sphériques, mais peuvent être légèrement ovales ou en forme de haricot. Les cocci peuvent être localisés isolément (microcoques); par paires (diplocoques); sous forme de chaînes (streptocoques) ou de grappes (staphylocoques), d'un paquet (sarcines). Les streptocoques peuvent provoquer une amygdalite et un érysipèle, des staphylocoques - divers processus inflammatoires et purulents.
bactérie en forme de bâtonnet le plus commun. Les bâtonnets peuvent être simples, reliés par paires (diplobactéries) ou en chaînes (streptobactéries). Les bactéries en forme de bâtonnet comprennent Escherichia coli, des agents pathogènes de la salmonellose, de la dysenterie, de la fièvre typhoïde, de la tuberculose, etc. Certaines bactéries en forme de bâtonnet ont la capacité de se former dans des conditions défavorables. des disputes. Les bâtonnets sporulés sont appelés bacilles. Les bacilles fusiformes sont appelés clostridies.
La sporulation est un processus complexe. Les spores diffèrent considérablement d'une cellule bactérienne normale. Ils ont une coquille dense et une très petite quantité d'eau, ils n'ont pas besoin de nutriments et la reproduction s'arrête complètement. Les spores sont capables de résister longtemps au dessèchement, aux hautes et basses températures et peuvent être dans un état viable pendant des dizaines et des centaines d'années (spores de charbon, botulisme, tétanos, etc.). Une fois dans un environnement favorable, les spores germent, c'est-à-dire qu'elles se transforment en la forme de propagation végétative habituelle.
Bactéries alambiquées peut être sous la forme d'une virgule - vibrions, avec plusieurs boucles - spirille, sous la forme d'un mince bâton torsadé - spirochètes. Les vibrions sont l'agent causal du choléra et l'agent causal de la syphilis est le spirochète.
9. Caractéristiques de la morphologie des rickettsies et des chlamydia
Les rickettsies sont de petits micro-organismes gram-négatifs caractérisés par un polymorphisme prononcé - ils forment des cocci, des formes en forme de bâtonnet et filamenteuses (Fig. 22). La taille des rickettsies varie de 0,5 à 3-4 microns, la longueur des formes filamenteuses atteint 10-40 microns. Ils ne forment pas de spores et de capsules, ils sont colorés en rouge selon Zdrodovsky.
Les chlamydia sont sphériques, ovoïdes ou en forme de bâtonnet. Leurs tailles fluctuent entre 0,2 et 1,5 microns. La morphologie et la taille des chlamydia dépendent du stade de leur cycle de développement intracellulaire, qui se caractérise par la transformation d'une petite formation élémentaire sphérique en un grand corps initial à division binaire. Avant de se diviser, les particules de chlamydia sont enveloppées dans une formation ressemblant à une capsule bactérienne. Coloration Chlamydia "* selon Romanovsky-Giemsa, gram-négatif, clairement visible dans les préparations intravitales avec microscopie à contraste de phase.
10. Structure et biologie des mycoplasmes.
Un seul ordre, Mycoplasmatales, appartient à la classe des Mollicutes. Représentants de cet ordre - mycoplasmes -
Ils diffèrent des bactéries en ce qu'ils n'ont pas de paroi cellulaire. Au lieu de cela, ils contiennent une membrane cytoplasmique de lipoprotéines à trois couches. La taille des mycoplasmes fluctue entre 125 et 250 microns. Ils sont ronds, ovales ou filamenteux, Gram-négatifs.
Les mycoplasmes se reproduisent par fission binaire, comme la plupart des bactéries, notamment après la formation de petites formations coccoïdes (corps élémentaires, EB) dans des structures filamenteuses.
Les mycoplasmes sont capables de bourgeonnement et de segmentation. L'unité de reproduction minimale est ET (0,7-0,2 microns). Le principal composant de la membrane cellulaire est le cholestérol. Les mycoplasmes ne sont pas capables de former du cholestérol et l'utilisent à partir de tissus ou de milieux nutritifs complétés par leur introduction. La coloration de Gram est négative, mais la coloration de Romanovsky-Giemsa donne les meilleurs résultats. Les mycoplasmes sont exigeants sur les conditions de culture : du sérum natif, du cholestérol, des acides nucléiques, des glucides, des vitamines et divers sels doivent être ajoutés aux milieux nutritifs. Sur les milieux denses, ils forment de petites colonies translucides caractéristiques au centre granuleux surélevé, leur donnant l'aspect d'"œufs au plat". Sur des milieux contenant du sang, certains types de mycoplasmes donnent une hémolyse a et bêta. Dans les milieux semi-liquides, les mycoplasmes se développent le long de la ligne d'injection, formant des colonies dispersées et friables. En milieu liquide, ils conduisent à un léger trouble ou opalescence ; certaines souches sont capables de former le film gras le plus fin. Chez l'homme, des représentants des genres Mycoplasma, Ureaplasma et Acholeplasma sont isolés, y compris des espèces pathogènes et saprophytes.