ფლაგელას მდებარეობის მიხედვით ბაქტერიები იყოფა. მოძრაობის ორგანოები: ფუნქციები და სტრუქტურა, პროტოზოების მოძრაობის მახასიათებლები
სტრუქტურა. ზედაპირზე არსებული ბაქტერიების ცნობილი სახეობების დაახლოებით ნახევარს აქვს მოძრაობის ორგანოები - ტალღისმაგვარი მრუდე ფლაგელა. ფლაგელას მასა ბაქტერიის მშრალი მასის 2%-მდეა. ფლაგელის სიგრძე მიკროორგანიზმის სხეულის სიგრძეზე მეტია და შეადგენს 3–12 მკმ; ფლაგელუმის სისქე 0,02 მკმ-ია, ხოლო პოლარული დროშები უფრო სქელია ვიდრე პერიტრიქული.
Flagella შედგება flagellin ცილისგან (ლათ. flagella - flagellum), რომელიც თავისი სტრუქტურით მიეკუთვნება მიოზინის ტიპის კონტრაქტურ ცილებს. ფლაგელუმი შეიცავს ან ერთ ერთგვაროვან ცილოვან ძაფს ან 2-3 ძაფს, რომლებიც მჭიდროდ არის დაკეცილი ლენტებით. ფლაგელუმის ძაფი არის ხისტი სპირალი, რომელიც გრეხილია საათის ისრის საწინააღმდეგოდ; სპირალის სიმაღლე სპეციფიკურია თითოეული ბაქტერიის სახეობისთვის.
დროშების რაოდენობა, ზომა და განლაგება არის მახასიათებლები, რომლებიც მუდმივია კონკრეტული სახეობისთვის და მხედველობაში მიიღება ტაქსონომიაში. თუმცა, ზოგიერთ ბაქტერიას შეუძლია ფლაგელის შექმნა. განსხვავებული ტიპები. გარდა ამისა, დროშების არსებობა დამოკიდებულია გარემო პირობებზე: გრძელვადიანი კულტივირების დროს ბაქტერიებს შეუძლიათ დაკარგონ დროშები, ხოლო თხევად გარემოზე მათი ხელახლა მიღება. ფლაგელას რაოდენობა და მდებარეობა იმავე სახეობაში შეიძლება განისაზღვროს სასიცოცხლო ციკლის სტადიით. ამიტომ, ამ მახასიათებლის ტაქსონომიური მნიშვნელობა არ უნდა იყოს გადაჭარბებული.
ბაქტერიების კლასიფიკაცია დროშების რაოდენობისა და ადგილმდებარეობის მიხედვით:
1. ატრიჩი - დროშები არ არის.
2. მონოტრიკუსი- უჯრედის ერთ-ერთ პოლუსზე მდებარე ერთი ფლაგელი (გვარი Vibrio)- ფლაგელას მონოპოლარული მონოტრიქული განლაგება, ყველაზე მოძრავი ბაქტერია.
3. პოლიტრიქუსი - ბევრი დროშები:
– ლოფოტრიხული- ფლაგელას შეკვრა უჯრედის ერთ ბოძზე (დაბადების ფსევდომონასი, ბურხოლდერია) - დროშების მონოპოლარული პოლიტრიქული განლაგება;
– ამფიტრიქული- უჯრედის თითოეულ პოლუსზე არის ფლაგელუმის შეკვრა (გვარი Spirillum)- დროშების ბიპოლარული პოლიტრიქული განლაგება;
– პერიტრიხული- დროშები განლაგებულია რაიმე განსაკუთრებული თანმიმდევრობით უჯრედის მთელ ზედაპირზე (ფამ. Enterobacteriaceae(დაბადების ეშერიხია, პროტეუსი), ფამ. bacillaceae,ოჯახი Clostidiaceae), დროშების რაოდენობა უჯრედში 6-დან 1000-მდე მერყეობს, ბაქტერიის ტიპის მიხედვით (სურ. 7).
ნახ.7.ბაქტერიებში flagella-ს ადგილმდებარეობის ვარიანტები:
1 - მონოტრიხი, 2 - ლოფოტრიხი;
3 - ამფიტრიქუსი; 4 - პერიტრიხი.
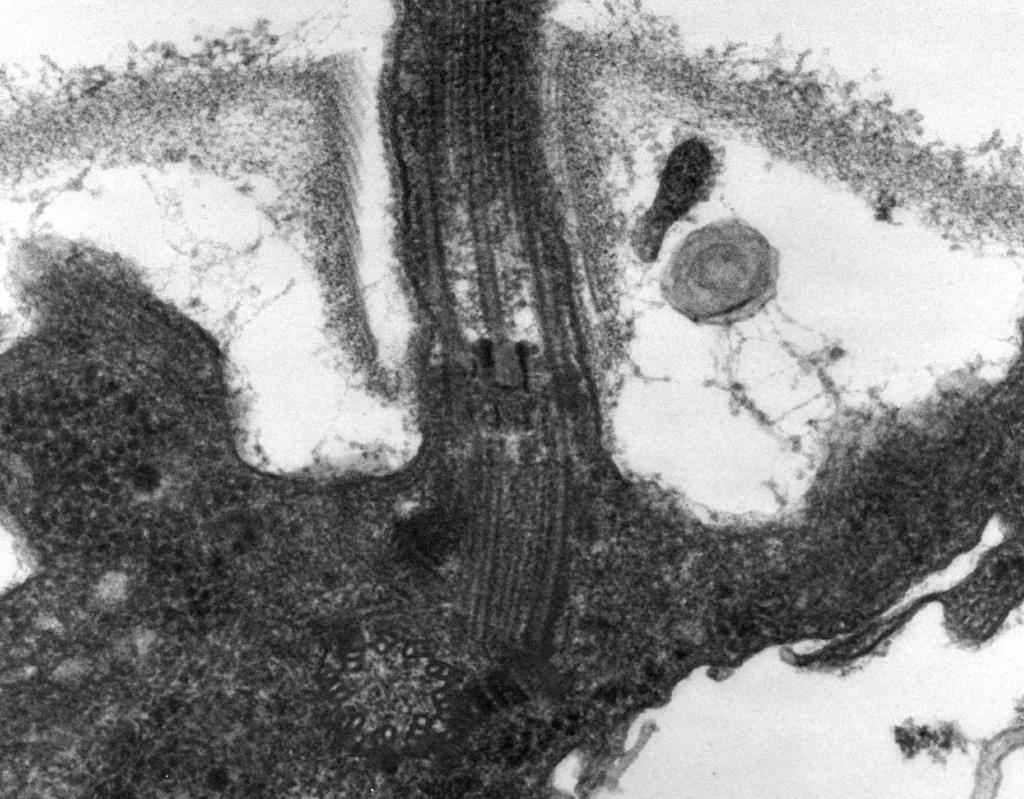
– ამას ცხადყოფს ელექტრონული მიკროსკოპია flagellum შედგება სამი ნაწილისაგან:სპირალური ძაფი, კაკალი და ბაზალური სხეული (სურ. 8).
ფლაგელის ძირითადი ნაწილი გრძელია სპირალური ძაფი (ფიბრილი) - ხისტი ღრუ ცილინდრი დაახლოებით 120 ნმ დიამეტრით, რომელიც შედგება ფლაგელინის ცილისგან. ცილის მოლეკულები ძაფის სიგრძეზე 11 რიგს ქმნიან და სპირალურადაა განლაგებული. ძაფის ზრდის დროს უჯრედის შიგნით სინთეზირებული ცილის მოლეკულები გადის ცილინდრის ღრუში და მიმაგრებულია მის ბოლოში სპირალზე. ფლაგელუმის ბოლოს არის ცილის თავსახური (სახურავი), რომელიც ხურავს ცილინდრის გახსნას და ხელს უშლის ცილის მოლეკულების გარემოში გათავისუფლებას. ფლაგელის ძაფის სიგრძემ შეიძლება მიაღწიოს რამდენიმე მიკრომეტრს. ბაქტერიების ზოგიერთ სახეობაში ფლაგელუმი დამატებით დაფარულია გარედან გარსით. CS-ის ზედაპირზე სპირალური ძაფი გადადის შესქელებულ მრუდე სტრუქტურაში - კაუჭში.

ბრინჯი. რვა.ფლაგელის სტრუქტურის სქემა
2. კაკალი(20-45 ნმ სისქის) უჯრედის ზედაპირთან არის შედარებით მოკლე ცილინდრი, რომელიც შედგება ფლაგელინისგან განსხვავებული ცილისგან და ემსახურება ძაფის მოქნილ კავშირს ბაზალურ სხეულთან.
3. ბაზალური სხეულიმდებარეობს ფლაგელის ძირში და უზრუნველყოფს მის ბრუნვას. ბაზალური სხეული შეიცავს 9-12 განსხვავებულ ცილას და შედგება ორი ან ოთხი დისკის (რგოლის)გან, რომელიც ღეროზეა დაკიდებული, რომელიც არის კაკლის გაგრძელება. ეს რგოლები დამონტაჟებულია CPM-სა და CS-ში. ორი შიდა რგოლი (M და S) არის ბაზალური სხეულის სავალდებულო კომპონენტები. M-რგოლი ლოკალიზებულია CPM-ში, S-რგოლი განლაგებულია გრამუარყოფითი ბაქტერიების პერიპლაზმურ სივრცეში ან გრამდადებითი ბაქტერიების პეპტიდოგლიკანურ პარკში. ორი გარე რგოლი (D და L) არ არის საჭირო მოძრაობისთვის, რადგან ისინი წარმოდგენილია მხოლოდ გრამუარყოფით ბაქტერიებში და ლოკალიზებულია, შესაბამისად, პეპტიდოგლიკანის შრეში და CL-ის გარე მემბრანაში. რგოლები S, D და L უმოძრაოა და ემსახურება ფლაგელის ფიქსაციას CS-ში. ფლაგელუმის ბრუნვა განისაზღვრება უჯრედის CPM-ში ჩაშენებული M-რგოლის ბრუნვით. ამრიგად, ფლაგელის ბაზალური სხეულის სტრუქტურული მახასიათებლები განისაზღვრება CS-ის სტრუქტურით.
ფუნქციურად, ბაზალური სხეული არის ელექტროძრავა, რომელიც იკვებება პროტონებით. ბაზალური სხეულის M-რგოლი (მბრუნავი როტორი) გარშემორტყმულია მემბრანის ცილებით უარყოფითი მუხტით (ძრავის სტატორი). ბაქტერიულ უჯრედს აქვს ელექტროქიმიური ენერგიის მექანიკურ ენერგიად გადაქცევის ეფექტური მექანიზმი. აქედან გამომდინარე, ბაქტერია ხარჯავს მთელი ენერგიის დაახლოებით 0,1%-ს, რომელსაც ის ხარჯავს ფლაგელის მუშაობაზე. ფლაგელუმის მოქმედების დროს გამოიყენება პროტონ-მოძრავი ძალა, რაც უზრუნველყოფილია მემბრანის გარე და შიდა მხარეებზე პროტონების კონცენტრაციის სხვაობით (გარედან უფრო მეტია) და მეტის არსებობით. უარყოფითი მუხტი მემბრანის შიდა მხარეს. პროტონის მამოძრავებელი ძალა აიძულებს პროტონებს ბაზალურ სხეულში გადავიდნენ უჯრედში, ხოლო ისინი ჩერდებიან როტორის გარკვეულ მონაკვეთებში და აძლევს მათ დადებით მუხტს, შემდეგ კი პროტონები შედიან უჯრედში. დამუხტული სექციები განლაგებულია ისე, რომ მიზიდულობის ძალა წარმოიქმნება როტორისა და სტატორის დამუხტულ მონაკვეთებს შორის, M- ბეჭედი იწყებს ბრუნვას დაახლოებით 300 ბრ/წთ სიჩქარით. ბრუნვის მექანიზმი: COOH ჯგუფის დამუხტვა-დატენვა ამინომჟავებში. ბეჭდის სრული ბრუნვისთვის ბაზალურ სხეულში 500-1000 პროტონმა უნდა გაიაროს. M- რგოლის ბრუნვა ღერძის მეშვეობით, რომელიც მყარად არის დაკავშირებული მასთან და კაკალი გადაეცემა ფლაგელის ძაფს, რომელიც ფუნქციონირებს როგორც პროპელერი ან გემის პროპელერი. ბაქტერია ცურავს მანამ, სანამ ხრახნი მუშაობს, ინერციის წვლილი უკიდურესად მცირეა.
გარდა ამისა, ბაქტერიები, თუნდაც მკვდარი, წყლის გარემოში ბრაუნის მოძრაობის შედეგად მოძრაობენ. ბაქტერიული უჯრედი მუდმივად ექვემდებარება გარემომცველი მოლეკულების ზემოქმედებას, რომლებიც თერმულ მოძრაობაში არიან. სხვადასხვა მიმართულებით დარტყმები ბაქტერიას გვერდიდან გვერდზე აგდებს.
ფლაგელას მოძრაობის ტიპი ბრუნვითია. არსებობს ორი სახის მოძრაობა: მართკუთხა და თაღლითური (პერიოდული შემთხვევითი ცვლილებები მოძრაობის მიმართულებით). როდესაც დროშები ბრუნავენ საათის ისრის საწინააღმდეგოდ (დაახლოებით 1 წამი), 40-60 ბრ/წთ სიხშირით (საშუალო ელექტროძრავის სიჩქარესთან ახლოს), მათი ძაფები იბნევა ერთ შეკვრაში (ნახ. 9a). ფლაგელას ბრუნვა გადაეცემა უჯრედს. ვინაიდან უჯრედი ფლაგელუმზე ბევრად მასიურია, ის იწყებს მოძრაობას სწორი ხაზით საპირისპირო მიმართულებით, 3-ჯერ ნაკლები სიჩქარით, ვიდრე ფლაგელუმის სიჩქარე.
ეს უზრუნველყოფს უჯრედის მთარგმნელობით მოძრაობას, რომლის სიჩქარეც თხევად გარემოშია განსხვავებული ტიპებიბაქტერია არის 20-200 მკმ/წმ (ეს შეესაბამება დაახლოებით 300-3000 სხეულის სიგრძეს წუთში) და უფრო ნელი მოძრაობა მყარი მედიის ზედაპირზე.
ბაქტერიას შეუძლია მიზანმიმართულად ცურვა ერთი მიმართულებით არაუმეტეს 3 წამის განმავლობაში, შემდეგ გარემომცველი მოლეკულების ზემოქმედება აქცევს მას შემთხვევითი მიმართულებით. ამიტომ ბაქტერიებმა შეიმუშავეს მოძრაობის მიმართულების სპონტანური ცვლილების მექანიზმი - ფლაგელარული ძრავის გადართვა. როდესაც ის იწყებს ბრუნვას საათის ისრის მიმართულებით (დაახლოებით 0,1 წმ), ბაქტერია ჩერდება და ტრიალდება (ახორციელებს „დალახვას“) შემთხვევითი მიმართულებით. ამ შემთხვევაში დროშები იფანტება სხვადასხვა მიმართულებით (სურ. 9ბ). ამფიტრიქოში გადაადგილებისას ფლაგელას ერთი შეკვრა შიგნიდან გარეთაა შემობრუნებული (როგორც ქარის მიერ შიგნიდან ამობრუნებული ქოლგა). შემდეგ ძრავა კვლავ გადადის საათის ისრის საწინააღმდეგო ბრუნვაზე და ბაქტერია ისევ ცურავს სწორი ხაზით, მაგრამ განსხვავებული, შემთხვევითი მიმართულებით.
ფლაგელას ასევე შეუძლია შეცვალოს მიმართულება გარე სტიმულის საპასუხოდ. თუ ბაქტერია მიიწევს მიზიდულობის ოპტიმალური კონცენტრაციისკენ, დროშები უჯრედს უბიძგებს გარემოში, მისი მართკუთხა მოძრაობა უფრო გრძელი ხდება და დახრის სიხშირე უფრო დაბალია, რაც საშუალებას აძლევს მას საბოლოოდ გადაადგილდეს სასურველი მიმართულებით.
ცნობილია არააქტიური (პარალიზებული) დროშების არსებობის შემთხვევები. ფლაგელირებული ბაქტერიების გადაადგილებისთვის აუცილებელია CS-ის მთლიანობა (დაუცველობა). უჯრედების დამუშავება ლიზოზიმით, რაც იწვევს CS-ის პეპტიდოგლიკანური ფენის მოცილებას, იწვევს ბაქტერიების გადაადგილების უნარის დაკარგვას, თუმცა დროშები ხელუხლებელი რჩება.
ბაქტერიების ტაქსი.Ნახვამდის გარემოუცვლელი რჩება, ბაქტერიები შემთხვევით ბანაობენ. თუმცა, გარემო იშვიათად არის სრულიად ერთგვაროვანი. თუ გარემო ჰეტეროგენულია, ბაქტერიები ავლენენ ელემენტარულ ქცევით რეაქციებს: ისინი აქტიურად მოძრაობენ გარკვეული გარე ფაქტორებით განსაზღვრული მიმართულებით. ბაქტერიების ასეთ გენეტიკურად განსაზღვრულ მიზანმიმართულ მოძრაობას ტაქსი ეწოდება. ფაქტორიდან გამომდინარე განასხვავებენ ქიმიოტაქსისს (განსაკუთრებული შემთხვევაა აეროტაქსისი), ფოტოტაქსისი, მაგნიტოტაქსისი, თერმოტაქსისი და ვისკოზიტაქსისი.
ქიმიოტაქსისი- მოძრაობა გარკვეული მიმართულებით ქიმიური ნივთიერების წყაროსთან შედარებით. ქიმიური ნივთიერებებიიყოფა ორ ჯგუფად: ინერტული და გამომწვევი ტაქსები - ქიმიოეფექტორები. ქიმიოეფექტორებს შორის არის ნივთიერებები, რომლებიც იზიდავს ბაქტერიებს - მიმზიდველებს (შაქარი, ამინომჟავები, ვიტამინები, ნუკლეოტიდები) და ნივთიერებები, რომლებიც მათ მოგერიებს - რეპელენტები (ზოგიერთი ამინომჟავა, ალკოჰოლი, ფენოლი, არაორგანული იონები). მიმზიდველი აერობული და ანაერობული პროკარიოტების მომგვრელი არის მოლეკულური ჟანგბადი. ატრაქტანტები ხშირად წარმოდგენილია საკვების სუბსტრატებით, თუმცა ორგანიზმისთვის აუცილებელი ყველა ნივთიერება არ მოქმედებს როგორც მიმზიდველი. ასევე, ყველა შხამიანი ნივთიერება არ არის რეპელენტური და ყველა საზიანო არ არის. ამრიგად, ბაქტერიებს არ შეუძლიათ რეაგირება რაიმე ნაერთზე, მაგრამ მხოლოდ გარკვეულ და განსხვავებულებს შეუძლიათ სხვადასხვა ბაქტერიისთვის.
ბაქტერიული უჯრედის ზედაპირულ სტრუქტურებში არის სპეციალური ცილის მოლეკულები - რეცეპტორები, რომლებიც სპეციალურად უკავშირდებიან გარკვეულ ქიმიოეფექტორს, ხოლო ქიმიოეფექტორის მოლეკულა არ იცვლება, მაგრამ კონფორმაციული ცვლილებები ხდება რეცეპტორის მოლეკულაში. რეცეპტორები განლაგებულია არათანაბრად უჯრედის მთელ ზედაპირზე და კონცენტრირებულია ერთ-ერთ პოლუსზე. რეცეპტორის მდგომარეობა ასახავს შესაბამისი ეფექტორის უჯრედგარე კონცენტრაციას.
ქიმიოტაქსის აქვს ადაპტაციური მნიშვნელობა. მაგალითად, ქოლერის ვიბრიოს ფორმები დაქვეითებული ქიმიოტაქსით ნაკლებად ვირუსულია.
აეროტაქსისი- ბაქტერიები, რომლებსაც მოლეკულური ჟანგბადი ესაჭიროებათ, გროვდება ჰაერის ბუშტუკების ირგვლივ, რომლებიც დაეცა საფარის ქვეშ.
ფოტოტაქსი- მოძრაობა სინათლისკენ ან მისგან მოშორებით, დამახასიათებელი ფოტოტროფული ბაქტერიებისთვის, რომლებიც იყენებენ სინათლეს ენერგიის წყაროდ.
მაგნიტოტაქსისი- წყლის ბაქტერიების უნარი, რომლებიც შეიცავს რკინის შემცველი მინერალების კრისტალებს, ცურავდნენ დედამიწის მაგნიტური ველის ხაზების გასწვრივ.
თერმოტაქსი- მოძრაობა ტემპერატურის ცვლილების მიმართულებით, რომელსაც აქვს დიდი მნიშვნელობაზოგიერთი პათოგენური ბაქტერიისთვის.
ვისკოზიტაქსისი- ხსნარის სიბლანტის ცვლილებებზე რეაგირების უნარი. ბაქტერიები ჩვეულებრივ მიდრეკილია უფრო მაღალი სიბლანტის მქონე გარემოსკენ, რაც დიდი მნიშვნელობა აქვს პათოგენური სახეობებისთვის.
მოცურების ბაქტერიები.დაბალი სიჩქარით (2-11 მკმ/წმ) სრიალის უნარი მყარ ან ბლანტ სუბსტრატზე აღმოჩენილია ზოგიერთ პროკარიოტში, მაგალითად, მიკოპლაზმაში.
არსებობს რამდენიმე ჰიპოთეზა, რომელიც ხსნის სრიალის მოძრაობას. Მიხედვით რეაქტიული ძრავის ჰიპოთეზაეს გამოწვეულია ლორწოს გამოყოფით სკ-ში ლორწოს მრავალრიცხოვანი ფორებით, რის შედეგადაც უჯრედი იხსნება სუბსტრატიდან ლორწოს გამოყოფის მიმართულების საპირისპირო მიმართულებით. Მიხედვით მოგზაურობის ტალღის ჰიპოთეზასრიალის მოძრაობა მოძრავი ფლაგელატებისგან თავისუფალ ფორმებში ასოცირდება პეპტიდოგლიკანის ფენასა და CL-ის გარე მემბრანას შორის მოწესრიგებული ფიბრილების თხელი ცილოვანი ფენის არსებობასთან, ფლაგელას ძაფების მსგავსი. ამ სტრუქტურების მიერ „გაშვებული“ ბოჭკოების ბრუნვითი მოძრაობა იწვევს უჯრედის ზედაპირზე „მოძრავი ტალღის“ (CL-ის მოძრავი მიკროსკოპული გამონაყარის) გამოჩენას, რის შედეგადაც უჯრედი იხრება სუბსტრატიდან. დაბოლოს, სტრუქტურები, რომლებიც წააგავს ფლიგელური ფორმების ბაზალურ სხეულებს, აღწერილია ზოგიერთ მოცურავ ბაქტერიაში.
ფლაგელას ფუნქციები:
1. უზრუნველყოს ადჰეზია - საწყისი ეტაპიინფექციური პროცესი.
2. უზრუნველყოს ბაქტერიების მოძრაობა.
3. განსაზღვრეთ ანტიგენური სპეციფიკა, ეს არის H-ანტიგენი.
დროშების გამოვლენა:
1. ნატურალური პრეპარატების ფაზის კონტრასტული მიკროსკოპია („დატეხილი“ და „დაკიდებული“ წვეთები). მიკროსკოპულად, მობილურობა განისაზღვრება ყოველდღიური კულტურის უჯრედებში. მობილურობის პასიური ბრაუნის მოძრაობისგან განასხვავების მიზნით, შესწავლილი კულტურის წვეთს ემატება ფენოლის 5%-იანი წყალხსნარის წვეთი; ამ შემთხვევაში აქტიური მოძრაობა ჩერდება.
2. ადგილობრივი პრეპარატების ბნელი ველის მიკროსკოპია.
3. საღებავებით ან ლითონებით შეღებილი პრეპარატების მსუბუქი მიკროსკოპია. ვინაიდან პრეპარატის მომზადებისას დროშები ძალიან ადვილად ზიანდება, ეს მეთოდები იშვიათად გამოიყენება ყოველდღიურ პრაქტიკაში.
დროშები შეღებილია აგარის დახრილობაზე გაზრდილი უჯრედების გამოყენებით. კონდენსაციის წყლის მახლობლად მდებარე უჯრედები შეირჩევა ბაქტერიული მარყუჟით და ფრთხილად გადადის იმავე ტემპერატურის სტერილურ გამოხდილ წყალში, როგორც ბაქტერიების ინკუბაციის ტემპერატურა დახრილ აგარს, და ბაქტერიები არ იშლება მარყუჟიდან, არამედ ფრთხილად ჩაეფლო წყალში. ბაქტერიით მილაკი ოთახის ტემპერატურაზე 30 წუთის განმავლობაში რჩება. გამოიყენება ქიმიურად სუფთა (ქრომის ნარევში გარეცხილი) მინა, რომელზედაც დაასხით 2-3 წვეთი სუსპენზია. სუსპენზია ვრცელდება მინის ზედაპირზე მისი ნაზად დახრილობით. გააშრეთ პრეპარატი ჰაერით.
დროშები ძალიან თხელია, ამიტომ მათი აღმოჩენა შესაძლებელია მხოლოდ სპეციალური დამუშავებით. თავდაპირველად აკრავის დახმარებით მიიღწევა შეშუპება და მათი ზომის მატება, შემდეგ კი ხდება პრეპარატის შეღებვა, რის გამოც ისინი ხილული ხდება სინათლის მიკროსკოპის ქვეშ.
უფრო ხშირად გამოიყენება ვერცხლის მეთოდი მოროზოვის მიხედვით (სურ. 10):
- პრეპარატი ფიქსირდება გამყინვარების ძმარმჟავას ხსნარით 1 წუთის განმავლობაში, გარეცხილია წყლით;
- წაისვით ტანინის ხსნარი (გარუჯვა, დროშების უფრო მკვრივი) 1 წუთის განმავლობაში, გარეცხილი წყლით;
- პრეპარატს ამუშავებენ ვერცხლის ნიტრატის გაჟღენთილი ხსნარით 1-2 წუთის განმავლობაში გახურებით, რეცხავენ წყლით, აშრობენ და ათვალიერებენ მიკროსკოპით.
მიკროსკოპულად, მუქი ყავისფერი უჯრედები და მსუბუქი დროშები ჩანს.

ბრინჯი. ათი.დროშების იდენტიფიცირება ვერცხლის საშუალებით
ბრინჯი. თერთმეტი.ფლაგელას იდენტიფიკაცია
ელექტრონული მიკროსკოპით
სურ. 4. მძიმე ლითონებით დეპონირებული პრეპარატების ელექტრონული მიკროსკოპია (სურ. 11).
5. ირიბად - ბაქტერიების ზრდის ბუნების მიხედვით, ნახევრად თხევად 0,3% აგარში დათესვისას. კულტურების თერმოსტატში 1-2 დღის განმავლობაში ინკუბაციის შემდეგ, აღინიშნება ბაქტერიების ზრდის ბუნება:
- არამოძრავ ბაქტერიებში (მაგ. S.saprophyticus) ინექციის გასწვრივ არის ზრდა - "ფრჩხილი", ხოლო საშუალო გამჭვირვალეა;
მოძრავ ბაქტერიებში (მაგ. E. coli) არის ზრდა ინექციის მხარეს, მთელ აგარის სვეტში - "ჰერინგბონი" და საშუალების დიფუზური სიმღვრივე.
ყველა ბაქტერიაიყოფა მობილურ და უძრავად. ბაქტერიებში მოძრაობის ორგანოებია flagella. ისინი შედგება ფლაგელინისგან, ცილისგან, რომელიც სტრუქტურულად დაკავშირებულია მიოზინის ტიპის კონტრაქტურ ცილებთან.
Flagellum ბაზაარის ბაზალური სხეული, რომელიც შედგება დისკების სისტემისგან (ბლეფაროპლასტი: 1 დისკი - უჯრედის კედლის გარე მხარე, 2 დისკი - უჯრედის კედლის შიდა მხარე, 3 დისკი - ციტოპლაზმური მემბრანა), "ჩაშენებული" ციტოპლაზმაში. მემბრანა და უჯრედის კედელი. ფლაგელის სიგრძე აღემატება თავად მიკრობის სხეულის სიგრძეს.
დროშების რაოდენობის მიხედვითდა მათი ადგილმდებარეობა, მოძრავი მიკროორგანიზმები იყოფა:
1. ერთფეროვანი, სხეულის ბოლოში ერთი ფლაგელი (ყველაზე მოძრავი). მაგალითად, Vibrio cholerae.
2. ლოფოტრიქოსი, უჯრედის ერთ-ერთ პოლუსზე ფლაგელას შეკვრა. მაგალითად, Burkholderia (Pseudomonas) pseudomalei არის მელიოიდოზის გამომწვევი აგენტი.
3. ამფიტრიქოსი, უჯრედის ორივე პოლუსზე ფლაგელუმის მქონე. მაგალითად, Spirillum volutans.
4. პერიტრიხული, უჯრედის მთელ პერიმეტრზე ფლაგელებით. მაგალითად, Escherichia coli, Salmonella typhi.
დროშების გამოვლენა.დროშები ძალიან თხელია, ამიტომ მათი აღმოჩენა შესაძლებელია მხოლოდ სპეციალური დამუშავებით. კერძოდ, ჯერ მორდანტის დახმარებით მიიღწევა შეშუპება და მათი ზომის მატება, შემდეგ კი ხდება პრეპარატის შეღებვა, რის გამოც ისინი ხილული ხდება სინათლის მიკროსკოპით. დროშების აღმოჩენა შესაძლებელია მოროზოვის, ლეფლერის მიხედვით შეღებვით, ასევე ელექტრონული მიკროსკოპით. Flagella ასევე შეიძლება გამოვლინდეს ბაქტერიების აქტიური მოძრაობა.
მიკრობების მოძრაობა შეინიშნება ცოცხალი კულტურებიდან „დაქუცმაცებული“ და „ჩამოკიდებული“ წვეთების პრეპარატებში. ეს პრეპარატები მიკროსკოპულია მშრალი ან ჩაძირული ლინზებით ბნელ ველში ან ფაზის კონტრასტში. გარდა ამისა, მოძრაობა შეიძლება განისაზღვროს ნახევრად თხევადი აგარში ბაქტერიების ზრდის ბუნებით.
დალიეს ბაქტერიებისგან.
Pili (pili), სინონიმები: villi, fimbria, არის ცილოვანი ბუნების თხელი ღრუ ძაფები, რომლებიც ფარავს ბაქტერიული უჯრედების ზედაპირს. flagella-სგან განსხვავებით, ისინი არ ასრულებენ საავტომობილო ფუნქციას.
Pili ვრცელდება უჯრედის ზედაპირიდან და შედგება ცილისგან პილინა.
მათი ფუნქციური დანიშნულების მიხედვით, ისინი იყოფა 2 ტიპად.
1) პირველი ტიპის პილი გვხვდება ბაქტერიების უმეტესობაში, ამიტომ მათ უწოდებენ "ზოგადი ტიპის ვილის" (ჩვეულებრივი პილი). ისინი იწვევენ ბაქტერიების მიმაგრებას ან ადჰეზიას მასპინძელი ორგანიზმის გარკვეულ უჯრედებთან. ადჰეზია არის ნებისმიერი ინფექციური პროცესის საწყისი ეტაპი.
2) მეორე ტიპის პილი (სინონიმები: კონიუგატიური, ან სექსუალური - სქესის პილი) ხელმისაწვდომია მხოლოდ დონორ ბაქტერიებში, რომლებსაც აქვთ სპეციალური პლაზმიდი. მათი რაოდენობა მცირეა - 1-4 უჯრედში.
იატაკის ხერხები ასრულებს შემდეგ ფუნქციებს:
1. ბაქტერიების კონიუგაციის დროს გენეტიკური მასალის ერთი უჯრედიდან მეორეში გადატანაში მონაწილეობა.
2. ადსორბირებენ სპეციფიკურ ბაქტერიულ ვირუსებს – ბაქტერიოფაგებს
ბაქტერიული სპორები, ფორმირების პირობები, მდებარეობა, მექანიზმი და აუჟესკის შეღებვის ეტაპები.
დაპირისპირება- მოსვენებული ბაქტერიების თავისებური ფორმა გრამდადებითი ტიპის უჯრედის კედლის სტრუქტურით.
სპორულაცია- ეს არის სახეობის (გენოფორის) შენარჩუნების გზა გარე გარემოში არახელსაყრელ პირობებში და არა გამრავლების მეთოდი.
წარმოიქმნება სპორებიბაქტერიების არსებობისთვის არახელსაყრელ პირობებში (გაშრობა, საკვები ნივთიერებების დეფიციტი და ა.შ.). ბაქტერიული უჯრედის შიგნით წარმოიქმნება ერთი სპორა (ენდოსპორი).
სპორულაციის ეტაპები
1. მოსამზადებელი. ბაქტერიების ციტოპლაზმაში იქმნება დატკეპნილი უბანი, რომელსაც არ აქვს თავისუფალი წყალი, რომელსაც ეწოდება „სპოროგენული ზონა“, რომელიც შეიცავს ნუკლეოიდს.
2. პრესპორების (პროსპორების) სტადია. სპოროგენული ზონის გარშემო წარმოიქმნება ორმაგი ციტოპლაზმური მემბრანის გარსი.
3. ქერქის ფორმირება, რომელიც შედგება პეპტიდოგლიკანისგან და კალციუმის მარილებისა და ლიპიდების მაღალი შემცველობით გარე გარსისგან.
4. მომწიფების ეტაპი. გარე გარსის გარე მხარეს წარმოიქმნება სპორის გარსი, რის შემდეგაც ხდება უჯრედის ვეგეტატიური ნაწილის ლიზირება, სპორის გათავისუფლება.
მიკრობიოლოგიის განვითარებამ მრავალი აღმოჩენა მოიტანა ბოლო ათწლეულების განმავლობაში. და ერთ-ერთი მათგანია ფლაგელირებული ბაქტერიების მოძრაობის თავისებურებები. ამ უძველესი ორგანიზმების ძრავების დიზაინი ძალიან რთული აღმოჩნდა და, მათი მუშაობის პრინციპის მიხედვით, ძალიან განსხვავდება პროტოზოების ჩვენი უახლოესი ევკარიოტული ნათესავების დროშებისგან. ფლაგელატის ბაქტერიის ძრავა ყველაზე მწვავე დაპირისპირება იყო კრეაციონისტებსა და ევოლუციონისტებს შორის. ბაქტერიების, მათი ფლაგელის ძრავების და მრავალი სხვა შესახებ - ეს სტატია.
ზოგადი ბიოლოგია
დასაწყისისთვის, გავიხსენოთ, როგორი ორგანიზმები არიან ისინი და რა ადგილი უჭირავთ ორგანული სამყაროს სისტემაში ჩვენს პლანეტაზე. ბაქტერიების დომენი აერთიანებს უამრავ უჯრედულ პროკარიოტულ (ფორმირებული ბირთვის გარეშე) ორგანიზმებს.
ეს ცოცხალი უჯრედები გამოჩნდნენ სიცოცხლის სცენაზე თითქმის 4 მილიარდი წლის წინ და იყვნენ პლანეტის პირველი დასახლებები. ისინი შეიძლება იყოს ძალიან განსხვავებული ფორმის (კოკები, წნელები, ვიბრიოები, სპიროქეტები), მაგრამ მათი უმეტესობა დროშებითაა.
სად ცხოვრობენ ბაქტერიები? Ყველგან. 5 × 10 30-ზე მეტი ცხოვრობს პლანეტაზე. დაახლოებით 40 მილიონი მათგანია 1 გრამ ნიადაგში, 39 ტრილიონამდე ცხოვრობს ჩვენს ორგანიზმში. ისინი გვხვდება მარიანას თხრილის ფსკერზე, ცხელ „შავ მწეველებში“ ოკეანეების ფსკერზე, ანტარქტიდის ყინულში და ამჟამად ხელებზე 10 მილიონამდე ბაქტერია გაქვთ.
ღირებულება უდაოა
მიუხედავად მათი მიკროსკოპული ზომისა (0,5-5 მიკრონი), მათი მთლიანი ბიომასა დედამიწაზე აღემატება ცხოველთა და მცენარეთა ბიომასას ერთად. მათი როლი ნივთიერებების მიმოქცევაში შეუცვლელია და მათი მომხმარებელთა (ორგანული ნივთიერებების დამღუპველი) თვისებები არ იძლევა საშუალებას, პლანეტა დაიფაროს გვამების მთებით.
ნუ დაივიწყებთ პათოგენებს: ჭირის, ჩუტყვავილას, სიფილისის, ტუბერკულოზის და მრავალი სხვა ინფექციური დაავადების გამომწვევი აგენტები ასევე ბაქტერიებია.
ბაქტერიებმა იპოვეს გამოყენება ადამიანის ეკონომიკურ საქმიანობაში. კვების მრეწველობისგან დაწყებული ( რძის პროდუქტებიყველი, მწნილი ბოსტნეული, ალკოჰოლური სასმელები), "მწვანე" ეკონომიკა (ბიოსაწვავი და ბიოგაზი) უჯრედების ინჟინერიის მეთოდებს და წამლების (ვაქცინები, შრატები, ჰორმონები, ვიტამინები) წარმოებას.

ზოგადი მორფოლოგია
როგორც უკვე აღვნიშნეთ, სიცოცხლის ამ ერთუჯრედულ წარმომადგენლებს არ აქვთ ბირთვი, მათი მემკვიდრეობითი მასალა (დნმ-ის მოლეკულები რგოლის სახით) მდებარეობს ციტოპლაზმის (ნუკლეოიდის) გარკვეულ არეალში. მათ უჯრედს აქვს პლაზმური მემბრანა და პეპტიდოგლიკან მურეინის მიერ წარმოქმნილი მკვრივი კაფსულა. უჯრედის ორგანელებიდან ბაქტერიებს აქვთ მიტოქონდრია, შეიძლება იყოს ქლოროპლასტები და სხვა სტრუქტურები სხვადასხვა ფუნქციით.
ბაქტერიების უმეტესობა არის დროშები. უჯრედის ზედაპირზე მჭიდრო კაფსულა ხელს უშლის მათ გადაადგილებას თავად უჯრედის შეცვლით, როგორც ამას აკეთებენ ამები. მათი flagella არის მკვრივი ცილის წარმონაქმნები სხვადასხვა სიგრძის და დაახლოებით 20 ნმ დიამეტრის. ზოგიერთ ბაქტერიას აქვს ერთი ფლაგელი (ერთფეროვანი), ზოგს კი ორი (ამფიტრიხული). ზოგჯერ დროშები განლაგებულია შეკვრაში (ლოფოტრიხული) ან ფარავს უჯრედის მთელ ზედაპირს (პერიტრიხულს).
ბევრი მათგანი ცხოვრობს როგორც ერთუჯრედიანი, მაგრამ ზოგიერთი ქმნის მტევანებს (წყვილები, ჯაჭვები, ძაფები, ჰიფები).

მოძრაობის მახასიათებლები
დროშის მქონე ბაქტერიებს შეუძლიათ სხვადასხვა გზით გადაადგილება. ზოგი მხოლოდ წინ მიიწევს და მიმართულებას ცვლის დახრის გზით. ზოგს შეუძლია კრუნჩხვა, ზოგს მოძრაობს სრიალით.
ბაქტერიების დროშები ასრულებენ არა მხოლოდ ფიჭური "ნიჩის" ფუნქციებს, არამედ შეიძლება იყოს "პანსიონის" ხელსაწყო.
ბოლო დრომდე ითვლებოდა, რომ ბაქტერიის ფლაგელი გველის კუდივით ქრიალებს. ბოლო კვლევებმა აჩვენა, რომ ბაქტერიების დროშა გაცილებით რთულია. მუშაობს როგორც ტურბინა. დისკზე მიმაგრებული, ის ბრუნავს ერთი მიმართულებით. ამძრავი, ანუ ბაქტერიების ფლაგელარული ძრავა, არის რთული მოლეკულური სტრუქტურა, რომელიც მუშაობს კუნთის მსგავსად. იმ განსხვავებით, რომ კუნთი უნდა მოდუნდეს შეკუმშვის შემდეგ და ბაქტერიული ძრავა მუდმივად მუშაობს.

ფლაგელის ნანომექანიზმი
მოძრაობის ბიოქიმიაში ჩაღრმავების გარეშე, ჩვენ აღვნიშნავთ, რომ ფლაგელის დრაივის შექმნაში მონაწილეობს 240-მდე ცილა, რომლებიც იყოფა 50 მოლეკულურ კომპონენტად, რომელსაც აქვს სისტემაში სპეციფიკური ფუნქცია.
ბაქტერიების ამ მამოძრავებელ სისტემაში არის როტორი, რომელიც მოძრაობს და სტატორი, რომელიც უზრუნველყოფს ამ მოძრაობას. არის ამძრავი ლილვი, ბუჩქი, გადაბმული, მუხრუჭები და ამაჩქარებლები
ეს მინიატურული ძრავა საშუალებას აძლევს ბაქტერიებს 35-ჯერ მეტი იმოგზაურონ საკუთარ ზომაზე მხოლოდ 1 წამში. ამავდროულად, თავად ფლაგელუმის მუშაობა, რომელიც წუთში 60 ათას ბრუნს აკეთებს, სხეული ხარჯავს მთელი ენერგიის მხოლოდ 0,1%-ს, რომელსაც უჯრედი მოიხმარს.
გასაკვირია ისიც, რომ ბაქტერიას შეუძლია შეცვალოს და შეაკეთოს თავისი საავტომობილო მექანიზმის ყველა სათადარიგო ნაწილი "გადაადგილებაში". უბრალოდ წარმოიდგინეთ, რომ თვითმფრინავში ხართ. და ტექნიკოსები ცვლიან მოძრავი ძრავის პირებს.
Flagellate ბაქტერია დარვინის წინააღმდეგ
ძრავა, რომელსაც შეუძლია იმუშაოს წუთში 60000 ბრუნვამდე სიჩქარით, თვითგაშვებით და საწვავად მხოლოდ ნახშირწყლების (შაქრის) გამოყენებით, ელექტროძრავის მსგავსი მოწყობილობის მქონე - შეიძლება თუ არა ასეთი მოწყობილობა წარმოშობილიყო ევოლუციის პროცესში?
ეს არის კითხვა, რომელიც მაიკლ ბეჰემ, ბიოლოგიის დოქტორმა, დაუსვა საკუთარ თავს 1988 წელს. მან ბიოლოგიაში შემოიტანა შეუქცევადი სისტემის ცნება - სისტემა, რომელშიც მისი ყველა ნაწილი ერთდროულად აუცილებელია მისი მუშაობის უზრუნველსაყოფად და ერთი ნაწილის მაინც ამოღება იწვევს მისი ფუნქციონირების სრულ დარღვევას.
დარვინის ევოლუციის თვალსაზრისით, სხეულში ყველა სტრუქტურული ცვლილება თანდათანობით ხდება და მხოლოდ წარმატებული შეირჩევა ბუნებრივი გადარჩევით.
მ.ბეჰეს დასკვნები, მოყვანილი წიგნში "დარვინის შავი ყუთი" (1996): ფლაგელირებული ბაქტერიის ძრავა არის 40-ზე მეტი ნაწილისგან შემდგარი განუყოფელი სისტემა და მინიმუმ ერთის არარსებობა გამოიწვევს მის სრულ დისფუნქციას. სისტემა, რაც ნიშნავს, რომ ეს სისტემა არ შეიძლებოდა მომხდარიყო ბუნებრივი გადარჩევის გზით.

ბალზამი კრეაციონისტებისთვის
შექმნის თეორიამ, რომელიც წარმოადგინა მეცნიერმა და ბიოლოგიის პროფესორმა, ბეთლემის ლეჰაი უნივერსიტეტის ბიოლოგიურ მეცნიერებათა ფაკულტეტის დეკანმა (აშშ) მ. ბეჰემ მაშინვე მიიპყრო ეკლესიის მსახურებისა და ღვთაებრივი თეორიის მომხრეების ყურადღება. სიცოცხლის წარმოშობა.
2005 წელს შეერთებულ შტატებშიც კი იყო სასამართლო პროცესი, სადაც ბეჰე იყო მოწმე "ინტელექტუალური დიზაინის" თეორიის მომხრეებისგან, რომელიც განიხილავდა კრეაციონიზმის შესწავლას დოვერის სკოლებში კურსში ". პანდაზე და ხალხზე“. პროცესი დაიკარგა, ასეთი საგნის სწავლება მოქმედი კონსტიტუციის საწინააღმდეგოდ იქნა აღიარებული.
მაგრამ დებატები კრეაციონისტებსა და ევოლუციონისტებს შორის დღესაც გრძელდება.
ფლაგელუმი არის ბაქტერიული უჯრედის ზედაპირული სტრუქტურა, რომელიც ემსახურება მათ თხევად მედიაში გადაადგილებისთვის.
ფლაგელას მდებარეობიდან გამომდინარე, ბაქტერიები იყოფა (ნახ. 1):
პერიტრიქიალური
შერეული
პოლუსი
სუბპოლარული
ბოძის დროშა- ერთი ან მეტი დროშა განლაგებულია უჯრედის ერთ (მონოპოლარულ) ან ორივე (ბიპოლარულ) პოლუსზე და ფუძე არის უჯრედის გრძელი ღერძის პარალელურად.
სუბპოლარული დროშები(სუბპოლარული) - ერთი ან მეტი ფლაგელა განლაგებულია უჯრედის გვერდითი ზედაპირის პოლუსზე გადასვლის ადგილას მის ერთ ან ორ ბოლოზე. ძირში არის სწორი კუთხე უჯრედის გრძელ ღერძთან.
გვერდითი დროშები(გვერდითი) - ერთი ან მეტი ფლაგელა შეკვრის სახით მდებარეობს უჯრედის ერთ-ერთი ნახევრის შუა წერტილში.
პერიტრიქიალური დროშები- განლაგებულია უჯრედის მთელ ზედაპირზე სათითაოდ ან ჩალიჩებით, ბოძები ჩვეულებრივ მოკლებულია მათ.
შერეული დროშები- ორი ან მეტი ფლაგელა მდებარეობს სხვადასხვა წერტილებიუჯრედები.
დროშების რაოდენობის მიხედვით გამოირჩევა:
მონოტრიხული - ერთი ფლაგელი
პოლიტრიხები - დროშების მტევანი
ასევე გამოირჩევა:
ლოფოტრიხული- დროშების მონოპოლიური პოლიტრიქიალური განლაგება.
ამფიტრიკები- დროშების ბიპოლარული პოლიტრიქიალური მოწყობა.
ბაქტერიული ფლაგელისა და ბაზალური სხეულის სტრუქტურა. Flagellum.
ფლაგელუმი განლაგებულია საკმაოდ მარტივად: ძაფი, რომელიც მიმაგრებულია ბაზალურ სხეულზე. ზოგჯერ ბაზალურ სხეულსა და ძაფს შორის შეიძლება ჩასვათ მილის მრუდი მონაკვეთი, ე.წ.
ქიმიური შემადგენლობით, ფლაგელუმი შედგება 98% ფლაგელინის ცილისგან (ფლაგელა - ფლაგელუმი), შეიცავს 16 ამინომჟავას, ჭარბობს გლუტამინი და ასპარტინის მჟავები, ტრიპტოფანი, ცისტეინი და ცისტინი არ არის მცირე რაოდენობით არომატულ ამინომჟავებში. ფლაგელინი აწესებს ანტიგენურ სპეციფიკას, მას H-ანტიგენი ეწოდება. ბაქტერიულ დროშებს არ გააჩნიათ ATPase აქტივობა.
ფლაგელის სისქე 10-12 ნმ, სიგრძე 3-15 მკმ.
ეს არის ხისტი სპირალი, გრეხილი საათის ისრის საწინააღმდეგოდ. ფლაგელუმის ბრუნვა ასევე ხორციელდება საათის ისრის საწინააღმდეგოდ, სიხშირით 40 rpm-დან 60 rpm-მდე, რაც იწვევს უჯრედის ბრუნვას საპირისპირო მიმართულებით, მაგრამ ვინაიდან უჯრედი გაცილებით მძიმეა ვიდრე ფლაგელი, მაშინ მისი ბრუნვა უფრო ნელია 12-დან 14 rpm-მდე.
Flagellum იზრდება დისტალური ბოლოდან, სადაც ქვედანაყოფები შედიან შიდა არხით. ზოგიერთ სახეობაში ფლაგელი გარედან დამატებით დაფარულია გარსით, რომელიც წარმოადგენს უჯრედის კედლის გაგრძელებას და, სავარაუდოდ, იგივე სტრუქტურა აქვს.
ბაზალური სხეული
ბაზალური სხეული შედგება 4 ნაწილისგან:
წნელების შეჯვარება ძაფით ან კაუჭით
ღეროზე დამაგრებული ორი დისკი. (M და S)
ცილის კომპლექსების ჯგუფი (სტატორები)
ცილის თავსახური
ბაქტერიებს, რომლებსაც აქვთ შიდა და გარე მემბრანა, აქვთ 2 დამატებითი დისკი (P და L) და ცილის სტრუქტურა, რომლებიც გარე მემბრანაზეა ბაზალური სხეულის მახლობლად, ამიტომ ისინი არ თამაშობენ მნიშვნელოვან როლს მოძრაობაში.
ბაზალური სხეულის სტრუქტურის თავისებურება განისაზღვრება უჯრედის კედლის სტრუქტურით: მისი ხელუხლებელი აუცილებელია ფლაგელას გადაადგილებისთვის. უჯრედების ლიზოზიმით დამუშავება იწვევს პეპტიდოგლიკანის შრის ამოღებას უჯრედის კედლიდან, რაც იწვევს მოძრაობის დაკარგვას, თუმცა ფლაგელის სტრუქტურა არ დაირღვა.
ბაქტერიული დროშები განსაზღვრავს ბაქტერიული უჯრედის მოძრაობას. Flagella არის თხელი ძაფები, რომლებიც წარმოიქმნება ციტოპლაზმური მემბრანიდან და უფრო გრძელია ვიდრე თავად უჯრედი. დროშები 12-20 ნმ სისქისა და 3-15 მკმ სიგრძისაა. ისინი შედგება 3 ნაწილისაგან: სპირალური ძაფი, კაკალი და ბაზალური სხეული, რომელიც შეიცავს ღეროს სპეციალური დისკებით (1 წყვილი დისკი გრამდადებითი და 2 წყვილი დისკი გრამუარყოფითი ბაქტერიებისთვის). ფლაგელას დისკები მიმაგრებულია ციტოპლაზმურ მემბრანასა და უჯრედის კედელზე. ეს ქმნის ელექტროძრავის ეფექტს საავტომობილო ჯოხით, რომელიც ბრუნავს ფლაგელუმს. Flagella შედგება ცილისგან - flagellin-ისგან (flagelum - flagellum); არის H ანტიგენი. ფლაგელინის ქვედანაყოფები დახვეულია. დროშების რაოდენობა სხვადასხვა სახეობის ბაქტერიებში მერყეობს ერთიდან (ერთფეროვანი) Vibrio cholerae-მდე ათ ან ასობით ფლაგელამდე, რომლებიც ვრცელდება ბაქტერიის (პერიტრიხის) პერიმეტრზე Escherichia coli-ში, Proteus-ში და ა.შ. უჯრედის დასასრული. ამფიტრიქოს აქვს ერთი ფლაგელი ან ფლაგელას შეკვრა უჯრედის მოპირდაპირე ბოლოებზე.
Pili (fimbriae, villi) - ძაფისებრი წარმონაქმნები, უფრო თხელი და მოკლე (3-10 ნმ x 0,3-10 მიკრონი), ვიდრე დროშები. პილი ვრცელდება უჯრედის ზედაპირიდან და შედგება პილინის ცილისგან, რომელსაც აქვს ანტიგენური მოქმედება. არსებობს პილი, რომელიც პასუხისმგებელია ადჰეზიაზე, ანუ დაზიანებულ უჯრედზე ბაქტერიების მიმაგრებაზე, ასევე კვებაზე, წყალ-მარილის მეტაბოლიზმზე და სექსუალურ (F-pili) ან კონიუგაციის პილიზე. სასმელი უხვად არის - გალიაში რამდენიმე ასეული. თუმცა, სქესობრივი პილი ჩვეულებრივ 1-3 უჯრედშია: ისინი წარმოიქმნება ეგრეთ წოდებული „მამაკაცის“ დონორის უჯრედებით, რომლებიც შეიცავს გადამდები პლაზმიდებს (F-, R-, კოლ-პლაზმიდები). სექს-პილის გამორჩეული თვისებაა ურთიერთქმედება სპეციალურ „მამრობით“ სფერულ ბაქტერიოფაგებთან, რომლებიც ინტენსიურად შეიწოვება სქესობრივ პილიზე.
სპორები არის მიძინებული ფირმისებრი ბაქტერიების თავისებური ფორმა, ე.ი. ბაქტერიები გრამდადებითი უჯრედის კედლის სტრუქტურით. სპორები წარმოიქმნება ბაქტერიების არსებობისთვის არახელსაყრელ პირობებში (გაშრობა, საკვები ნივთიერებების დეფიციტი და ა.შ.. ბაქტერიის უჯრედში წარმოიქმნება ერთი სპორა (ენდოსპორი), სპორების წარმოქმნა ხელს უწყობს სახეობების შენარჩუნებას და არ წარმოადგენს გამრავლების მეთოდს. სოკოების მსგავსად. Bacillus გვარის სპორის წარმომქმნელ ბაქტერიებს აქვთ სპორები, რომლებიც არ აღემატება უჯრედის დიამეტრს. ბაქტერიებს, რომელთა სპორის ზომა აღემატება უჯრედის დიამეტრს, ეწოდება კლოსტრიდიუმი, მაგალითად, Clostridium გვარის ბაქტერიებს (ლათ. Clostridium - spindle). სპორები მჟავა რეზისტენტულია, ამიტომ წითლად შეღებილია აუჯესკის მეთოდით ან ზიელ-ნილსენის მეთოდით, ხოლო ვეგეტატიური უჯრედი ლურჯად.
დავის ფორმა შეიძლება იყოს ოვალური, სფერული; საკანში მდებარეობა ტერმინალურია, ე.ი. ჯოხის ბოლოს (ტეტანუსის გამომწვევში), სუბტერმინალური - ჯოხის ბოლოსთან უფრო ახლოს (ბოტულიზმის, გაზის განგრენის პათოგენებში) და ცენტრალური (ჯილეხის ბაცილებში). სპორა დიდხანს ნარჩუნდება მრავალშრიანი გარსის, კალციუმის დიპიკოლინატის, დაბალი წყლის შემცველობის და დუნე მეტაბოლური პროცესების არსებობის გამო. ხელსაყრელ პირობებში სპორები აღმოცენდება სამი თანმიმდევრული სტადიით: გააქტიურება, დაწყება, გაღივება.
8. ბაქტერიების ძირითადი ფორმები
გლობულური ბაქტერიები (კოკები)ჩვეულებრივ სფერული, მაგრამ შეიძლება იყოს ოდნავ ოვალური ან ლობიოს ფორმის. კოკები შეიძლება განთავსდეს ცალკე (მიკროკოკები); წყვილებში (დიპლოკოკები); ჯაჭვების (სტრეპტოკოკები) ან ყურძნის მტევნების (სტაფილოკოკები) სახით, შეფუთვა (სარცინები). სტრეპტოკოკმა შეიძლება გამოიწვიოს ტონზილიტი და ერიზიპელა, სტაფილოკოკები - სხვადასხვა ანთებითი და ჩირქოვანი პროცესები.
ღეროს ფორმის ბაქტერიებიყველაზე გავრცელებული. წნელები შეიძლება იყოს ერთჯერადი, დაკავშირებული წყვილებში (დიპლობაქტერიები) ან ჯაჭვებში (სტრეპტობაქტერიები). ღეროს ფორმის ბაქტერიებს მიეკუთვნება Escherichia coli, სალმონელოზის პათოგენები, დიზენტერია, ტიფური ცხელება, ტუბერკულოზი და ა.შ. დავები.სპორების წარმომქმნელი ღეროები ე.წ ბაცილი.შუბლისებრი ბაცილები ე.წ კლოსტრიდია.
სპორულაცია რთული პროცესია. სპორები მნიშვნელოვნად განსხვავდება ნორმალური ბაქტერიული უჯრედისგან. მათ აქვთ მკვრივი გარსი და ძალიან მცირე რაოდენობით წყალი, არ საჭიროებენ საკვებ ნივთიერებებს და რეპროდუქცია მთლიანად ჩერდება. სპორები უძლებენ გაშრობას, მაღალ და დაბალ ტემპერატურას დიდი ხნის განმავლობაში და შეიძლება სიცოცხლისუნარიან მდგომარეობაში იყვნენ ათეულობით და ასობით წლის განმავლობაში (ჯილეხის სპორები, ბოტულიზმი, ტეტანუსი და ა.შ.). ხელსაყრელ გარემოში მოხვედრის შემდეგ სპორები აღმოცენდებიან, ანუ გადაიქცევიან ჩვეულებრივ ვეგეტატიურ გამრავლებად ფორმაში.
ჩახლართული ბაქტერიაშეიძლება იყოს მძიმის სახით - ვიბრიოები, რამდენიმე ხვეულით - სპირილა, წვრილი გრეხილი ჯოხის სახით - სპიროქეტები. ვიბრიოები არის ქოლერის გამომწვევი აგენტი, ხოლო სიფილისის გამომწვევი არის სპიროქეტა.
9. რიკეტსიისა და ქლამიდიის მორფოლოგიის თავისებურებები
რიკეტზია მცირე გრამუარყოფითი მიკროორგანიზმებია, რომლებსაც ახასიათებთ გამოხატული პოლიმორფიზმი - ისინი ქმნიან კოკებს, ღეროსებრ და ძაფისებრ ფორმებს (სურ. 22). რიკეტსიის ზომები მერყეობს 0,5-დან 3-4 მიკრონიმდე, ძაფისებრი ფორმების სიგრძე 10-40 მიკრონს აღწევს. ისინი არ წარმოქმნიან სპორებს და კაფსულებს, ისინი წითლად შეღებილია ზდროდოვსკის მიხედვით.
ქლამიდია არის სფერული, ოვალური ან ღეროს ფორმის. მათი ზომები მერყეობს 0,2-1,5 მიკრონის ფარგლებში. ქლამიდიის მორფოლოგია და ზომა დამოკიდებულია მათი უჯრედშორისი განვითარების ციკლის სტადიაზე, რომელიც ხასიათდება მცირე სფერული ელემენტარული წარმონაქმნის ტრანსფორმირებით დიდ საწყის სხეულად ბინარული გაყოფით. გაყოფამდე ქლამიდიის ნაწილაკები ბაქტერიული კაფსულის მსგავსი წარმონაქმნით არის მოცული. ქლამიდიის ლაქა "* რომანოვსკი-გიემსას მიხედვით, გრამუარყოფითი, აშკარად ჩანს ინტრავიტალურ პრეპარატებში ფაზა-კონტრასტული მიკროსკოპით.
10. მიკოპლაზმების სტრუქტურა და ბიოლოგია.
მხოლოდ ერთი ორდენი, Mycoplasmatales, ეკუთვნის Mollicutes კლასს. ამ ორდენის წარმომადგენლები - მიკოპლაზმები -
ისინი განსხვავდებიან ბაქტერიებისგან იმით, რომ მათ არ აქვთ უჯრედის კედელი. ამის ნაცვლად, ისინი შეიცავს სამ ფენის ლიპოპროტეინების ციტოპლაზმურ მემბრანას. მიკოპლაზმების ზომები მერყეობს 125-250 მიკრონის ფარგლებში. ისინი მრგვალი, ოვალური ან ძაფისებრია, გრამუარყოფითი.
მიკოპლაზმები მრავლდება ორობითი დაშლით, ისევე როგორც ბაქტერიების უმეტესობა, განსაკუთრებით ძაფისებრ სტრუქტურებში მცირე კოკოიდური წარმონაქმნების (ელემენტარული სხეულები, EB) წარმოქმნის შემდეგ.
მიკოპლაზმას შეუძლია კვირტის და სეგმენტაციის უნარი. მინიმალური გამრავლების ერთეულია ET (0,7-0,2 მიკრონი). უჯრედის მემბრანის მთავარი კომპონენტია ქოლესტერინი. მიკოპლაზმებს არ შეუძლიათ ქოლესტერინის ფორმირება და გამოიყენონ იგი ქსოვილებიდან ან მკვებავი მედიებიდან, რომლებიც დამატებულია მათი შეყვანით. გრამ-შეღებვა უარყოფითია, მაგრამ რომანოვსკი-გიემსას შეღებვა იძლევა საუკეთესო შედეგს. მიკოპლაზმები მოითხოვს კულტივირების პირობებს: მკვებავ გარემოს უნდა დაემატოს მშობლიური შრატი, ქოლესტერინი, ნუკლეინის მჟავები, ნახშირწყლები, ვიტამინები და სხვადასხვა მარილები. მკვრივ მედიაზე ისინი ქმნიან დამახასიათებელ მცირე გამჭვირვალე კოლონიებს აწეული მარცვლოვანი ცენტრით, რაც მათ "შემწვარი კვერცხების" იერს აძლევს. სისხლთან ერთად მიკოპლაზმების ზოგიერთი ტიპი იძლევა ა- და ბეტა ჰემოლიზს. ნახევრად თხევად გარემოში, მიკოპლაზმები იზრდება ინექციის ხაზის გასწვრივ, ქმნიან დისპერსიულ, დამსხვრეულ კოლონიებს. თხევად გარემოში, ისინი იწვევენ მცირე ნისლს ან ოფლიანობას; ზოგიერთ შტამს შეუძლია შექმნას ყველაზე თხელი ცხიმიანი ფილმი. ადამიანებში იზოლირებულია Mycoplasma, Ureaplasma და Acholeplasma გვარების წარმომადგენლები, მათ შორის პათოგენური და საპროფიტული სახეობები.