べん毛の位置に応じて、細菌はに分けられます。 運動のオルガネラ:機能と構造、原生動物の運動の特徴
構造。 表面にいる細菌の既知の種の約半分は、波のように湾曲したべん毛という運動器官を持っています。 べん毛の質量は、細菌の乾燥質量の最大 2% です。 鞭毛の長さは微生物の体の長さよりも長く、3 ~ 12 μm です。 鞭毛の厚さは0.02μmで、極鞭毛は周毛よりも厚いです。
べん毛はフラジェリンタンパク質(ラテン語鞭毛 - 鞭毛)で構成されており、その構造はミオシン型の収縮タンパク質に属しています。 べん毛には、1 本の均一なタンパク質フィラメントまたは 2 ~ 3 本のフィラメントがしっかりと折りたたまれて編組されています。 鞭毛の糸は、反時計回りにねじれた硬いらせんです。 らせんのピッチは、各細菌種に固有です。
鞭毛の数、サイズ、および配置は、特定の種で一定の特徴であり、分類学で考慮されます。 ただし、一部の細菌は鞭毛を形成することができます。 他の種類. さらに、鞭毛の存在は環境条件に依存します。長期培養中の固体培地では、細菌は鞭毛を失う可能性があり、液体培地では再び獲得される可能性があります。 同じ種の鞭毛の数と位置は、生活環の段階によって決定できます。 したがって、この特徴の分類学的重要性を過大評価すべきではありません。
べん毛の数と位置による細菌の分類:
1.アトリチ -鞭毛はありません。
2.モノトリクス- 細胞の極の 1 つに位置する 1 つの鞭毛 (ビブリオ属)- 最も移動性の高いバクテリアである鞭毛の単極単毛配置。
3. Polytrichus - 多くのべん毛:
– ろくでなし- 細胞の一方の極にあるべん毛の束 (誕生 シュードモナス, バークホルデリア) - べん毛の単極多毛配置。
– 両生類- 細胞の各極には鞭毛の束があります (スピリラム属)- 鞭毛のバイポーラ多毛配置;
– 周産期- 鞭毛は、細胞の表面全体に特定の順序で配置されていません (fam. 腸内細菌科(誕生 エシェリヒア、プロテウス)、ファム。 バチルス科、家族 アブラナ科)、鞭毛の数は、細菌の種類に応じて、細胞あたり 6 から 1000 までさまざまです (図 7)。
図7。細菌の鞭毛の位置のオプション:
1 - モノリッチ、2 - ロフォトリッチ。
3 - 両生類; 4 - ペリトリク。
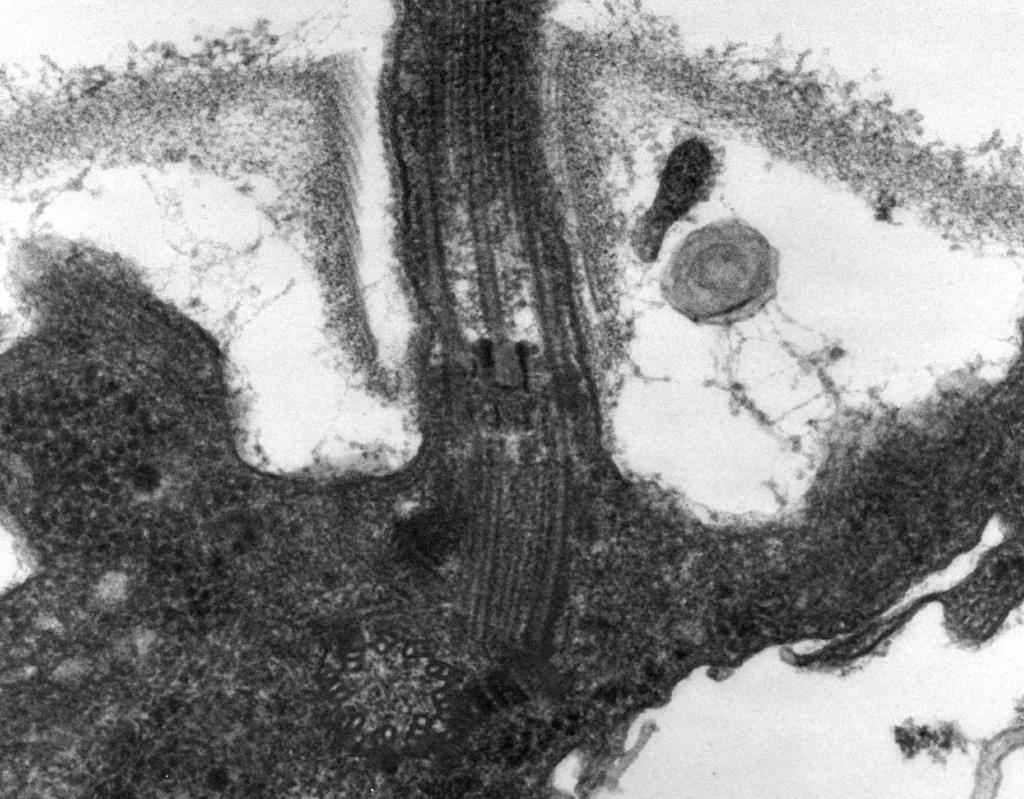
– 電子顕微鏡で明らかになったこと 鞭毛は 3 つの部分で構成されています。らせんフィラメント、フック、および基底体 (図 8)。
鞭毛の主要部分は長い スパイラルスレッド (フィブリル) - フラジェリンタンパク質からなる直径約 120 nm の堅い中空シリンダー。 タンパク質分子は、糸の長さに沿って 11 列を形成し、らせん状に配置されています。 糸が伸びる過程で、細胞内で合成されたタンパク質分子がシリンダーの空洞を通過し、その端でらせん状にくっつきます。 鞭毛の端には、シリンダーの開口部を閉じて環境へのタンパク質分子の放出を防ぐタンパク質キャップ (蓋) があります。 鞭毛フィラメントの長さは数マイクロメートルに達することがあります。 細菌のいくつかの種では、鞭毛はさらに外側が鞘で覆われています。 CSの表面では、らせん状の糸が肥厚した湾曲した構造、つまりフックに入ります。

米。 8。鞭毛の構造のスキーム
2.フック(厚さ 20 ~ 45 nm) は、細胞表面近くの比較的短いシリンダーで、フラジェリンとは異なるタンパク質で構成されており、基底体へのフィラメントの柔軟な接続を提供するのに役立ちます。
3. 基礎体鞭毛の基部に位置し、その回転を保証します。 基底体には 9 ~ 12 の異なるタンパク質が含まれており、フックの続きであるロッドに張られた 2 つまたは 4 つのディスク (リング) で構成されています。 これらのリングは、CPM と CS に取り付けられています。 2 つの内側のリング (M と S) は、基底体の必須コンポーネントです。 MリングはCPMに局在し、Sリングはグラム陰性菌のペリプラズム空間またはグラム陽性菌のペプチドグリカン嚢に位置しています。 2 つの外側のリング (D と L) は、グラム陰性菌にのみ存在し、それぞれペプチドグリカン層と CL の外膜に局在しているため、移動にはオプションです。 リング S、D、および L は不動であり、CS の鞭毛を固定するのに役立ちます。 鞭毛の回転は、セルの CPM に組み込まれた M リングの回転によって決まります。 したがって、鞭毛の基底体の構造的特徴は、CSの構造によって決定されます。
機能的には、基底体は陽子によって駆動される電気モーターです。 基底体の M リング (回転するローター) は、負電荷を持つ膜タンパク質 (モーター ステーター) に囲まれています。 細菌細胞には、電気化学エネルギーを機械エネルギーに変換する効率的なメカニズムがあります。 したがって、バクテリアは、消費する全エネルギーの約 0.1% を鞭毛の操作に費やします。 鞭毛が機能するとき、プロトン駆動力が使用されます。これは、膜の外側と内側のプロトン濃度の違い (外側の方が多い) と、より負の電荷の存在によって提供されます。膜の内側にあります。 陽子の原動力により、陽子は基底体を通過して細胞内に入りますが、陽子はローターの特定の部分にとどまり、陽電荷を与えて細胞内に入ります。 帯電部分は、ローターとステーターの帯電部分の間に引力が発生し、Mリングが約300 rpmの速度で回転し始めるように配置されています。 回転メカニズム: アミノ酸の COOH 基の充電-再充電。 リングが完全に 1 回転するには、500 ~ 1000 個の陽子が基底体を通過する必要があります。 Mリングにしっかりと接続された軸とフックを介したMリングの回転は、プロペラまたは船のプロペラとして機能する鞭毛フィラメントに伝達されます。 細菌はスクリューが作動している限り泳ぎますが、慣性の寄与は非常に小さいです。
さらに、水生環境のバクテリアは、死んだものでさえ、ブラウン運動の結果として移動します。 細菌細胞は、熱運動をしている周囲の分子の影響を常に受けています。 さまざまな方向からのヒットは、バクテリアを左右に投げます。
鞭毛の動きのタイプは回転です。 動きには、直線とタンブリング (動きの方向が周期的にランダムに変化する) の 2 種類があります。 べん毛が反時計回り (約 1 秒) で 40 ~ 60 rpm (平均的な電気モーターの速度に近い) の速度で回転すると、それらの糸は 1 つの束に織り込まれます (図 9a)。 べん毛の回転が細胞に伝わります。 細胞は鞭毛よりもはるかに重いため、鞭毛の速度の 3 分の 1 の速度で反対方向に直線的に動き始めます。
これにより、細胞の並進運動が保証され、その速度は液体培地で 他の種類バクテリアは 20 ~ 200 μm/秒 (これは 1 分あたり約 300 ~ 3000 体の長さに相当します) であり、固体培地の表面では動きが遅くなります。
バクテリアは意図的に一方向に 3 秒以上泳ぐことができず、周囲の分子の衝撃によりランダムな方向に向きを変えます。 したがって、バクテリアは、運動方向の自発的な変化 - べん毛モーターの切り替え - のメカニズムを開発しました。 時計回りに回転し始めると (約 0.1 秒)、バクテリアは停止し、ランダムな方向に反転します (「タンブル」を実行します)。 この場合、べん毛はさまざまな方向に散らばります(図9b)。 両生類では、移動するとき、鞭毛の 1 つの束が裏返されます (傘が風によって裏返されるように)。 次に、モーターが再び反時計回りに切り替わり、バクテリアは再び直線的に泳ぎますが、方向はランダムです。
べん毛は、外部刺激に反応して方向を変えることもできます。 細菌が最適な誘引物質濃度に向かって移動すると、鞭毛が細胞を培地に押し込み、その直線運動が長くなり、タンブリング頻度が低くなり、最終的に目的の方向に移動できるようになります。
不活性な(麻痺した)べん毛が存在する場合が知られています。 鞭毛細菌の移動には、CS の完全性 (完全性) が必要です。 リゾチームで細胞を処理すると、CS のペプチドグリカン層が除去され、バクテリアの移動能力が失われますが、鞭毛はそのまま残ります。
バクテリアのタクシー。さよなら 環境バクテリアはランダムに泳ぎます。 ただし、環境が完全に均一であることはめったにありません。 環境が不均一な場合、細菌は基本的な行動反応を示します。細菌は、特定の外的要因によって決定される方向に積極的に移動します。 このような遺伝的に決定されたバクテリアの意図的な動きは、タクシーと呼ばれます。 因子に応じて、走化性(特別な場合は気流走性)、走光性、走磁性、熱走性および粘性走性が区別されます。
走化性- 化学物質の供給源に対する特定の方向への動き。 化学物質 2つのグループに分けられます:不活性および原因となるタクシー - 化学作用因子。 化学作用物質の中には、バクテリアを誘引する物質 - 誘引物質 (糖、アミノ酸、ビタミン、ヌクレオチド) と、バクテリアを寄せ付けない物質 - 忌避剤 (一部のアミノ酸、アルコール、フェノール、無機イオン) があります。 好気性原核生物の好気性誘引剤と嫌気性原核生物の忌避剤は分子状酸素です。 誘引物質は、多くの場合、食品基質によって表されますが、体に必要なすべての物質が誘引物質として機能するわけではありません。 また、すべての有毒物質が忌避剤として機能するわけではなく、すべての忌避剤が有害であるとは限りません。 したがって、細菌はどの化合物にも反応できず、細菌ごとに特定の異なる化合物にのみ反応します。
細菌細胞の表面構造には、特定の化学エフェクターに特異的に結合する受容体である特別なタンパク質分子がありますが、化学エフェクター分子は変化しませんが、受容体分子で立体構造の変化が起こります。 受容体は細胞の表面全体に不均一に配置されており、極の 1 つに集中しています。 受容体の状態は、対応するエフェクターの細胞外濃度を反映しています。
走化性には適応値があります。 例えば、走化性が損なわれた形態のコレラビブリオは毒性が低い。
エアロタキシス- 酸素分子を必要とする細菌は、カバースリップの下に落ちた気泡の周りに蓄積します。
走光性- 光をエネルギー源として使用する光合成バクテリアの特徴である、光に近づく、または遠ざかる動き。
磁気走性- 鉄を含む鉱物の結晶を含む水生細菌が地球の磁場の線に沿って泳ぐ能力。
熱走性- 温度変化の方向への動き。 非常に重要一部の病原菌に。
粘性持続性- 溶液の粘度の変化に対応する能力。 細菌は通常、粘性の高い培地を好む傾向があり、これは病原性種にとって非常に重要です。
滑走菌。固体または粘性の基質上を低速 (2 ~ 11 µm/s) でスライドする能力は、マイコプラズマなどの一部の原核生物で発見されています。
スライド運動を説明するいくつかの仮説があります。 によると ジェット推進仮説これは、SC の多数の粘液孔を介した粘液の分泌によるものであり、その結果、細胞は粘液分泌の方向とは反対の方向に基質からはじかれます。 によると 進行波仮説鞭毛を含まない運動性形態での滑走運動は、鞭毛のフィラメントと同様に、規則正しい原繊維の薄いタンパク質層のペプチドグリカン層と CL の外膜との間の存在に関連しています。 これらの構造によって「起動」されたフィブリルの回転運動は、細胞表面に「進行波」(CLの微視的な膨らみの移動)の出現をもたらし、その結果、細胞は基質から反発されます。 最後に、鞭毛のある形態の基底体に似た構造が、いくつかの滑空細菌で説明されています。
べん毛の機能:
1. 接着性を付与する - 初期感染プロセス。
2.細菌の運動性を提供します。
3. 抗原特異性を決定します。これが H 抗原です。
べん毛の検出:
1. ネイティブ製剤の位相差顕微鏡 (「粉砕」および「吊り下げ」ドロップ)。 微視的に、可動性は毎日の培養細胞で決定されます。 可動性と受動的なブラウン運動を区別するために、フェノールの 5% 水溶液を研究対象の培養液に 1 滴加えると、能動的な動きが停止します。
2. ネイティブ製剤の暗視野顕微鏡検査。
3. 染料または金属で染色された標本の光学顕微鏡検査。 鞭毛は準備の準備中に非常に簡単に損傷するため、これらの方法は日常の練習ではめったに使用されません。
べん毛は、寒天斜面で成長した細胞を使用して染色されます。 凝縮水の近くにある細胞をバクテリア ループで選択し、傾斜寒天上でのバクテリアの培養温度と同じ温度の滅菌蒸留水に慎重に移し、バクテリアをループから振り落とさずに注意深く水に浸します。 細菌の入ったチューブを室温で 30 分間放置します。 化学的に純粋な(クロム混合物で洗浄された)ガラスが使用され、その上に2〜3滴の懸濁液が適用されます。 ゆっくりと傾けて懸濁液をガラス面に広げます。 薬剤を自然乾燥させます。
鞭毛は非常に薄いため、特別な処理をしないと検出できません。 最初に、エッチングの助けを借りて、膨潤とサイズの増加が達成され、次に準備が着色され、光学顕微鏡で見えるようになります。
より頻繁に使用される モロゾフによる銀メッキ法 (図 10):
- 調製物を氷酢酸溶液で 1 分間固定し、水で洗浄する。
- タンニン溶液(なめし、べん毛をより密にする)を1分間塗布し、水で洗います。
-調製物は、硝酸銀の含浸溶液で1~2分間加熱することにより処理され、水で洗浄され、乾燥され、顕微鏡検査される。
顕微鏡では、暗褐色の細胞と明るいべん毛が見えます。

米。 十。シルバリングによるべん毛の同定
米。 十一。べん毛の同定
電子顕微鏡による
図 4. 重金属を堆積させた標本の電子顕微鏡写真 (図 11)。
5. 間接的 - 半液体 0.3% 寒天に播種した場合の細菌増殖の性質による。 作物をサーモスタットで 1 ~ 2 日間培養した後、細菌の増殖の性質が記録されます。
- 非運動性細菌 (例えば. S.saprophyticus)注射に沿って成長があります-「爪」、そして媒体は透明です;
運動性細菌(例えば. 大腸菌) 寒天カラム - 「ヘリンボーン」、および培地のびまん性濁度全体を通して、注射の側に成長があります。
すべての細菌可動式と不動式に分けられます。 細菌の運動器官は鞭毛です。 それらは、構造的にミオシン型収縮タンパク質に関連するタンパク質であるフラジェリンで構成されています。
べん毛基部ディスクのシステムからなる基底体です(眼瞼形成体:1ディスク - 細胞壁の外側、2ディスク - 細胞壁の内側、3ディスク - 細胞質膜)、細胞質に「埋め込まれています」膜と細胞壁。 鞭毛の長さは、微生物自体の体の長さよりも長くなります。
べん毛の数によるとおよびそれらの場所、運動性微生物は次のように分類されます。
1. 体の端に鞭毛が 1 つある (最も可動性が高い)。 たとえば、コレラ菌。
2. Lophotrichous、細胞の極の 1 つに鞭毛の束があります。 たとえば、バークホルデリア (シュードモナス) シュードマレイは類鼻疽の原因物質です。
3. 両生類で、細胞の両極に鞭毛があります。 たとえば、Spirillum volutans。
4. 細胞の周囲全体にべん毛がある、周毛性。 例えば、大腸菌、チフス菌。
べん毛の検出。鞭毛は非常に薄いため、特別な処理をしないと検出できません。 特に、最初に、媒染剤の助けを借りて、膨潤とサイズの増加が達成され、次に製剤が着色されるため、光学顕微鏡で見えるようになります。 べん毛は、モロゾフ、レフラーによる染色、および電子顕微鏡によって検出できます。 べん毛は、細菌の活発な運動性によっても検出できます。
微生物の動きは、生きた培養物からの「つぶれた」滴と「ぶら下がった」滴の準備で観察されます。 これらの標本は、暗視野または位相コントラストでドライレンズまたは液浸レンズを使用して顕微鏡で観察されます。 さらに、運動性は、半液体寒天における細菌の増殖の性質によって決定することができます。
バクテリアから飲む。
ピリ(線毛)、同義語:絨毛、線毛は、細菌細胞の表面を覆うタンパク質性の細い中空フィラメントです。 べん毛とは異なり、それらは運動機能を実行しません。
線毛は細胞表面から伸び、タンパク質で構成されています ピリナ.
それらの機能目的によると、それらは2つのタイプに分けられます。
1) 第一型の線毛は、ほとんどの細菌に存在するため、「一般型絨毛」(一般線毛)と呼ばれます。 それらは、宿主生物の特定の細胞への細菌の付着または接着を引き起こします。 接着は、感染プロセスの初期段階です。
2) 2 番目のタイプの線毛 (類義語: 接合性、または性的線毛) は、特別なプラスミドを持つドナー細菌でのみ利用できます。 それらの数は少なく、セルあたり 1 ~ 4 個です。
フロアソーは次の機能を果たします。
1. 細菌の接合中に、ある細胞から別の細胞への遺伝物質の移動に参加します。
2. 特定の細菌ウイルスを吸着する - バクテリオファージ
Aujeszky 染色の細菌胞子、形成条件、位置、メカニズムおよび段階。
論争- グラム陽性型の細胞壁構造を持つ、特異な形態の休止細菌。
胞子形成- これは、悪条件の下で種 (ジェノフォア) を外部環境で保存する方法であり、繁殖の方法ではありません。
胞子が形成される細菌の存在に不利な条件下(乾燥、栄養不足など)。 細菌の細胞内には、1つの胞子(エンドスポア)が形成されます。
胞子形成の段階
1.準備。 細菌の細胞質では、核様体を含む「胞子形成帯」と呼ばれる、自由水を持たない圧縮された領域が形成されます。
2.前胞子(前胞子)の段階。 胞子形成帯の周囲には、二重の細胞質膜の鞘が形成されます。
3. ペプチドグリカンとカルシウム塩と脂質を多く含む外膜からなる皮質の形成。
4.成熟の段階。 外膜の外側に胞子エンベロープが形成された後、細胞の栄養部分が溶解され、胞子が放出されます。
微生物学の発展は、ここ数十年で多くの発見をもたらしました。 そしてその一つが鞭毛細菌の動きの特異性です。 これらの古代生物のエンジンの設計は非常に複雑であることが判明し、その作業の原理によれば、原生動物に最も近い真核生物の親戚の鞭毛とは大きく異なります。 鞭毛細菌のエンジンは、創造論者と進化論者の間で最も熱い論争となっています。 バクテリア、そのべん毛モーターなどについて - この記事。
一般生物学
まず、それらがどのような生物であり、地球上の有機世界のシステムでどのような場所を占めているかを思い出してください。 バクテリア ドメインは、膨大な数の単細胞原核生物 (形成された核を持たない) を結合します。
これらの生きた細胞は、ほぼ 40 億年前に生命の舞台に登場し、地球の最初の入植者でした。 それらは非常に異なる形状(球菌、桿菌、ビブリオ、スピロヘータ)である可能性がありますが、それらのほとんどは鞭毛です.
細菌はどこに住んでいますか? どこにでも。 地球上には 5 × 10 30 以上が生息しています。 1グラムの土壌には約4000万個、私たちの体には最大39兆個が住んでいます. それらは、マリアナ海溝の底、海底のホットな「ブラックスモーカー」、南極大陸の氷の中にあり、現在、手には最大1,000万個のバクテリアがいます.
価値は間違いない
それらの微視的なサイズ (0.5 ~ 5 ミクロン) にもかかわらず、地球上のそれらの総バイオマスは、動物と植物のバイオマスを合わせたものよりも大きくなります。 物質の循環におけるそれらの役割はかけがえのないものであり、消費者(有機物の破壊者)の特性により、地球が死体の山で覆われることはありません。
さて、病原体を忘れないでください。ペスト、天然痘、梅毒、結核、および他の多くの感染症の原因物質も細菌です。
細菌は、人間の経済活動に応用されています。 食品業界から( 乳製品、チーズ、ピクルス、 アルコール飲料)、「グリーン」経済(バイオ燃料とバイオガス)から細胞工学の方法と医薬品(ワクチン、血清、ホルモン、ビタミン)の生産まで。

一般的な形態
すでに述べたように、これらの単細胞生命体には核がなく、それらの遺伝物質(リング状のDNA分子)は細胞質(核様体)の特定の領域に位置しています。 彼らの細胞は、原形質膜と、ペプチドグリカンムレインによって形成された高密度のカプセルを持っています。 細胞小器官のうち、細菌にはミトコンドリアがあり、葉緑体やさまざまな機能を持つ他の構造が存在する場合があります。
ほとんどの細菌は鞭毛です。 細胞の表面にあるタイトなカプセルは、アメーバが行うように、細胞自体を変化させることによってそれらが動き回るのを防ぎます. それらの鞭毛は、さまざまな長さと直径約 20 nm の密なタンパク質形成です。 鞭毛を 1 つ持つ細菌 (単毛類) もあれば、鞭毛を 2 つ持つ細菌 (両生類) もあります。 鞭毛が束になって配置されている場合(葉状毛)、または細胞の表面全体を覆っている場合(毛周)があります。
それらの多くは単一細胞として生きていますが、一部はクラスター (ペア、鎖、フィラメント、菌糸) を形成します。

動きの特徴
鞭毛細菌はさまざまな方法で移動できます。 前にしか動かず、タンブリングで方向を変えるものもある。 けいれんできるものもあれば、スライドして動くものもあります。
バクテリアのべん毛は、細胞の「オール」の機能を果たすだけでなく、「搭乗」ツールにもなります。
最近まで、バクテリアの鞭毛はヘビの尾のように振ると信じられていました。 最近の研究は、細菌の鞭毛がはるかに複雑であることを示しています。 タービンのように機能します。 ドライブに取り付けられ、一方向に回転します。 アクチュエーター、または細菌のべん毛モーターは、筋肉のように機能する複雑な分子構造です。 筋肉は収縮後に弛緩しなければならず、バクテリアのモーターは常に働いているという違いがあります.

鞭毛のナノメカニズム
動きの生化学を掘り下げることなく、最大 240 のタンパク質が鞭毛ドライブの作成に関与していることに注目します。これらのタンパク質は、システム内で特定の機能を持つ 50 の分子コンポーネントに分割されます。
このバクテリアの推進システムには、動くローターと、この動きを提供するステーターがあります。 ドライブシャフト、ブッシング、クラッチ、ブレーキ、アクセルがあります
この小型のエンジンにより、バクテリアはわずか 1 秒間で自身のサイズの 35 倍移動することができます。 同時に、毎分6万回転する鞭毛自体の働きは、細胞が消費する全エネルギーのわずか0.1%しか消費しません。
また驚くべきことに、バクテリアはそのモーター機構のすべてのスペアパーツを「外出先で」交換および修理できます。 あなたが飛行機に乗っていると想像してみてください。 また、技術者は稼働中のモーターのブレードを交換します。
鞭毛細菌対ダーウィン
毎分 60,000 回転までの速度で作動し、自己始動し、燃料として炭水化物 (砂糖) のみを使用し、電気モーターに似た装置を備えたエンジン - そのような装置は進化の過程で生じたでしょうか?
これは、生物学の博士号を持つマイケル・ベーエが 1988 年に自問した質問です。 彼は、還元不可能なシステムの概念を生物学に導入しました。これは、その機能を確実にするためにすべての部分が同時に必要であり、少なくとも1つの部分を取り除くとその機能が完全に破壊されるシステムです。
ダーウィンの進化の観点から、身体の構造変化はすべて徐々に起こり、成功したものだけが自然淘汰によって選択されます。
本「ダーウィンのブラック ボックス」(1996 年) で述べられている M. Behe の結論: べん毛細菌のエンジンは 40 を超える部分からなる不可分なシステムであり、少なくとも 1 つが欠けていると、細菌の完全な機能不全につながります。システム、つまり、このシステムは自然淘汰によって発生した可能性はありません。

創造論者のためのバーム
ベツレヘムのリーハイ大学(米国)の生物科学学部長である科学者であり生物学の教授であるM. Beheによって提示された創造の理論は、すぐに教会の牧師と神の理論の支持者の注目を集めました生命の起源。
2005年、米国では、コースのドーバーの学校での創造論の研究の導入を考慮した「合理的なデザイン」の理論の支持者から、ベヘが証人となった訴訟さえありました」パンダと人について。」 プロセスが失われ、そのような主題の教えは現在の憲法に反するものとして認識されました。
しかし、創造論者と進化論者の間の議論は今日も続いています。
鞭毛は細菌細胞の表面構造であり、液体培地での移動に役立ちます。
鞭毛の位置に応じて、細菌は次のように分類されます (図 1)。
周膜
混合
ポール
サブポーラー
極鞭毛- 1 つまたは複数の鞭毛が細胞の一方 (単極) または両方 (双極) の極に位置し、基部が細胞の長軸に平行である。
亜極性鞭毛(亜極) - 1つまたは複数の鞭毛が、その端の1つまたは2つで、細胞の極への外側面の移行点に位置しています。 基部では、セルの長軸と直角です。
側鞭毛(外側) - 束の形をした 1 つまたは複数の鞭毛が、細胞の半分の 1 つの中間点にあります。
周毛鞭毛- セルの表面全体に1つずつまたは束になって配置されているため、通常、極はそれらを奪われています。
混合鞭毛- 鞭毛が 2 つ以上ある 異なる点細胞。
鞭毛の数に応じて、次のものがあります。
Monotrichous - 1 つの鞭毛
ポリトリクス - べん毛の束
また、区別されます:
ろくでなし- べん毛の多毛性配置を独占する。
両生類- べん毛のバイポーラ多毛性配置。
細菌の鞭毛と基底体の構造。 べん毛。
鞭毛自体は非常に単純に配置されています: 基底体に取り付けられたフィラメントです。 チューブの曲がった部分、いわゆるフックを基底体とフィラメントの間に挿入できる場合があります。これはフィラメントよりも太く、基底体へのフィラメントの柔軟な取り付けに関与しています。
化学組成に関しては、鞭毛は98%のフラジェリンタンパク質(鞭毛-鞭毛)で構成され、16個のアミノ酸が含まれ、グルタミンとアスパラギン酸が優勢で、トリプトファン、システイン、シスチンは少量の芳香族アミノ酸に含まれていません。 フラジェリンは抗原特異性を課し、H 抗原と呼ばれます。 細菌のべん毛には ATPase 活性がありません。
鞭毛の厚さは 10 ~ 12 nm、長さは 3 ~ 15 μm です。
それは、反時計回りにねじれた硬いらせんです。 鞭毛の回転も反時計回りに 40 rpm から 60 rpm の頻度で行われ、これにより細胞は逆方向に回転しますが、 セルは鞭毛よりもはるかに重いため、その回転は12〜14 rpmと遅くなります。
鞭毛は、サブユニットが内部チャネルを通って入る遠位端から成長します。 一部の種では、鞭毛の外側がさらに鞘で覆われています。これは細胞壁の続きであり、おそらく同じ構造を持っています。
基礎体
基底体は 4 つの部分で構成されています。
フィラメントまたはフックとのロッド嵌合
棒につながれた2枚の円盤。 (MとS)
タンパク質複合体のグループ (固定子)
プロテインキャップ
内膜と外膜を持つ細菌は、基底体近くの外膜にある 2 つの追加のディスク (P と L) とタンパク質構造を持っているため、それらは運動に重要な役割を果たしません。
基底体の構造の特異性は、細胞壁の構造によって決まります。鞭毛の動きには、その無傷が必要です。 リゾチームで細胞を処理すると、細胞壁からペプチドグリカン層が除去され、動きが失われますが、鞭毛の構造は乱されませんでした。
細菌のべん毛は、細菌細胞の運動性を決定します。 べん毛は、細胞質膜に由来する細いフィラメントで、細胞自体よりも長い。 べん毛の厚さは 12 ~ 20 nm、長さは 3 ~ 15 µm です。 それらは 3 つの部分で構成されています: らせん状の糸、フック、および特別なディスク (グラム陽性菌用の 1 対のディスクとグラム陰性菌用の 2 対のディスク) を備えたロッドを含む基部本体。 べん毛のディスクは、細胞質膜と細胞壁に付着しています。 これにより、鞭毛を回転させるモーター ロッドを備えた電気モーターの効果が生まれます。 鞭毛は、タンパク質 - フラジェリン(鞭毛 - 鞭毛から)で構成されています。 H抗原です。 フラジェリンサブユニットはコイル状になっています。 さまざまな種の細菌の鞭毛の数は、コレラ菌の 1 つ (単毛) から、大腸菌、プロテウスなどの細菌の周囲に沿って伸びる 10 または数百の鞭毛 (ペリトリク) までさまざまです。セルの終わり。 両生類には、細胞の両端に 1 つの鞭毛または鞭毛の束があります。
線毛 (線毛、絨毛) - 鞭毛よりも細くて短い (3 ~ 10 nm x 0.3 ~ 10 ミクロン) の糸状体。 線毛は細胞表面から伸びており、抗原活性を持つ線毛タンパク質で構成されています。 接着を担う線毛、つまり影響を受けた細胞に細菌を付着させる線毛と、栄養、水-塩代謝および性的 (F-線毛)、または抱合線毛を担う線毛があります。 飲み物はたくさんあります - ケージごとに数百。 ただし、性線毛は通常、細胞あたり 1 ~ 3 です。性線毛は、伝達可能なプラスミド (F、R、Col プラスミド) を含むいわゆる「男性」ドナー細胞によって形成されます。 性線毛の際立った特徴は、性線毛に集中的に吸着される特別な「雄」の球状バクテリオファージとの相互作用です。
胞子は、休眠中のファーミキュート バクテリアの特異な形態です。 グラム陽性の細胞壁構造を持つ細菌。 胞子は、細菌の存在にとって好ましくない条件下(乾燥、栄養不足など)で形成されます。1つの胞子(エンドスポア)は細菌細胞の内部で形成されます。胞子の形成は種の保存に寄与し、繁殖の方法ではありません, 真菌のように. バチルス属の胞子形成細菌は、細胞の直径を超えない胞子を持っています. 胞子のサイズが細胞の直径を超える細菌は、クロストリジウムと呼ばれます, 例えば、クロストリジウム属の細菌 (lat.クロストリジウム - スピンドル)。胞子は耐酸性であるため、Aujeszky法またはZiehl-Neelsen法に従って赤色に染色され、栄養細胞は青色に染色されます.
紛争の形は、楕円形、球形の場合があります。 セル内の位置はターミナル、つまり スティックの端(破傷風の原因物質)、サブターミナル - スティックの端に近い(ボツリヌス中毒、ガス壊疽の病原体)および中央(炭疽菌)。 胞子は、多層シェル、ジピコリン酸カルシウム、低水分含有量、および代謝プロセスの遅さの存在により、長期間持続します。 好条件の下では、胞子は活性化、開始、発芽の 3 つの段階を経て発芽します。
8. 細菌の主な形態
球状細菌(球菌)通常は球形ですが、わずかに楕円形または豆の形をしている場合があります。 球菌は単独で見つけることができます(小球菌)。 ペアで(双球菌); チェーン (レンサ球菌) またはブドウの房 (ブドウ球菌)、パッケージ (サルシナス) の形で。 連鎖球菌は、扁桃炎および丹毒、ブドウ球菌 - さまざまな炎症性および化膿性のプロセスを引き起こす可能性があります。
棒状菌最も一般的な。 桿菌は、単一で、ペアで接続されている場合もあれば (ディプロバクテリア)、鎖状である場合もあります (ストレプトバクテリア)。 棒状菌には、大腸菌、サルモネラ症、赤痢、腸チフス、結核などの病原体が含まれます。一部の棒状菌は、悪条件下で形成される能力を持っています。 論争。胞子形成棒は呼ばれます 細菌。紡錘状桿菌と呼ばれる クロストリジウム。
胞子形成は複雑なプロセスです。 胞子は、通常の細菌細胞とは大きく異なります。 それらは密な殻と非常に少量の水を持ち、栄養素を必要とせず、繁殖は完全に停止します。 胞子は、乾燥、高温および低温に長時間耐えることができ、数十年から数百年にわたって生存可能な状態を保つことができます (炭疽菌、ボツリヌス菌、破傷風などの胞子)。 好ましい環境になると、胞子は発芽します。つまり、胞子は通常の栄養繁殖形態に変わります。
複雑な細菌コンマ - ビブリオ、いくつかのカール - スピリラ、細いねじれた棒 - スピロヘータの形をとることができます。 ビブリオはコレラの原因物質であり、梅毒の原因物質はスピロヘータです。
9. リケッチアとクラミジアの形態の特徴
リケッチアは、顕著な多形性を特徴とする小さなグラム陰性微生物であり、球菌、棒状および糸状の形態を形成します(図22)。 リケッチアのサイズは 0.5 ~ 3 ~ 4 ミクロンで、繊維状の長さは 10 ~ 40 ミクロンに達します。 それらは胞子やカプセルを形成せず、Zdrodovskyによると赤く染色されています.
クラミジアは、球形、卵形、または棒状です。 それらのサイズは 0.2 ~ 1.5 ミクロン以内で変動します。 クラミジアの形態とサイズは、細胞内発生サイクルの段階に依存します。これは、小さな球状の基本的な形成が二分裂を伴う大きな最初の体に変化することを特徴としています。 分裂する前に、クラミジア粒子は細菌の莢膜に似た構造に包まれています。 クラミジア染色「* Romanovsky-Giemsa によると、グラム陰性で、位相差顕微鏡による生体内製剤ではっきりと見えます。
10. マイコプラズマの構造と生物学。
モリクテス綱に属する目はマイコプラズマ目だけです。 このオーダーの代表 - マイコプラズマ -
それらは細胞壁がないという点でバクテリアとは異なります。 代わりに、3 層のリポタンパク質細胞質膜が含まれています。 マイコプラズマのサイズは、125 ~ 250 ミクロンの範囲で変動します。 それらは円形、楕円形または糸状で、グラム陰性です。
マイコプラズマは、ほとんどの細菌と同様に、特に糸状構造で小さなコッコイド形成 (素体、EB) が形成された後、二分裂によって繁殖します。
マイコプラズマは出芽と分節が可能です。 最小再生単位は ET (0.7 ~ 0.2 ミクロン) です。 細胞膜の主成分はコレステロールです。 マイコプラズマはコレステロールを形成することができず、導入された組織または栄養培地からコレステロールを利用します。 それらはグラムによると陰性に染色されますが、ロマノフスキー-ギムザ染色が最良の結果をもたらします。 マイコプラズマは培養条件に厳しい条件を課しています。天然の血清、コレステロール、核酸、炭水化物、ビタミン、およびさまざまな塩を栄養培地に追加する必要があります。 密度の高い培地では、中心部が盛り上がった粒状の特徴的な小さな半透明のコロニーを形成し、「目玉焼き」のように見えます。 血液を含む培養液では、ある種のマイコプラズマがα溶血とβ溶血を起こします。 半液体培地では、マイコプラズマは注入ラインに沿って成長し、分散したもろいコロニーを形成します。 液体媒体では、それらはわずかな曇りまたは乳白色を引き起こします。 いくつかの株は、最も薄い脂っこいフィルムを形成することができます. ヒトでは、マイコプラズマ属、ウレアプラズマ属、アコレプラズマ属の代表が分離されており、病原性および腐生性種が含まれています。