Ըստ դրոշակի տեղակայման՝ բակտերիաները բաժանվում են. Շարժման օրգաններ. ֆունկցիաներ և կառուցվածք, նախակենդանիների շարժման առանձնահատկությունները
Կառուցվածք. Մակերեւույթի վրա գտնվող բակտերիաների հայտնի տեսակների մոտ կեսն ունի շարժման օրգաններ՝ ալիքանման կոր դրոշակ: Դրոշակի զանգվածը մանրէի չոր զանգվածի մինչև 2%-ն է։ Դրոշակի երկարությունն ավելի երկար է, քան միկրոօրգանիզմի մարմնի երկարությունը և կազմում է 3–12 մկմ; դրոշակի հաստությունը 0,02 մկմ է, իսկ բևեռային դրոշակները ավելի հաստ են, քան պերիտրիկները։
Դրոշակները կազմված են flagellin սպիտակուցից (լատիներեն flagella - flagellum), որն իր կառուցվածքով պատկանում է միոզին տեսակի կծկվող սպիտակուցներին։ Դրոշակը պարունակում է կա՛մ մեկ միատարր սպիտակուցային թել, կա՛մ 2-3 թելեր, որոնք սերտորեն ծալված են հյուսի մեջ: Դրոշակի թելը կոշտ պարույր է, որը պտտվում է ժամացույցի սլաքի ուղղությամբ. խխունջի բարձրությունը հատուկ է յուրաքանչյուր բակտերիաների տեսակի համար:
Դրոշակների քանակը, չափը և դասավորությունը որոշակի տեսակի համար հաստատուն հատկանիշներ են և հաշվի են առնվում տաքսոնոմիայում: Այնուամենայնիվ, որոշ բակտերիաներ կարող են ձևավորել դրոշակ: տարբեր տեսակներ. Բացի այդ, դրոշակի առկայությունը կախված է շրջակա միջավայրի պայմաններից. երկարատև աճեցման ընթացքում բակտերիաները կարող են կորցնել դրոշակները, իսկ հեղուկ միջավայրում դրանք կարող են նորից ձեռք բերել: Դրոշակների քանակը և գտնվելու վայրը նույն տեսակի մեջ կարող է որոշվել կյանքի ցիկլի փուլով: Հետեւաբար, այս հատկանիշի տաքսոնոմիկ նշանակությունը չպետք է գերագնահատել:
Բակտերիաների դասակարգումը ըստ դրոշակների քանակի և տեղակայման.
1. Ատրիչի - դրոշակները բացակայում են:
2. Մոնոտրիխուս- մեկ դրոշակ, որը գտնվում է բջիջի բևեռներից մեկում (սեռ Vibrio)- Դրոշակների՝ ամենաշարժական բակտերիաների միաբևեռ միաձույլ դասավորությունը:
3. Polytrichus - շատ դրոշակներ.
– լոֆոտրիկ- բջջի մեկ բևեռում դրոշակների փաթեթ (ծնունդ Pseudomonas, Բուրկհոլդերիա) - Դրոշակների միաբևեռ պոլիտրիկ դասավորություն;
– ամֆիտրիկ- բջջի յուրաքանչյուր բևեռում դրոշակի մի կապ կա (սեռ Spirillum)- դրոշակների երկբևեռ պոլիտրիկ դասավորություն;
– peritrichous- Դրոշակները դասավորված են առանց որոշակի հերթականության բջջի ամբողջ մակերեսի վրա (ֆամ. Enterobacteriaceae(ծնունդ Էշերիխիա, Պրոտեուս), հանր. bacillaceae,ընտանիք Clostidiaceae), դրոշակների թիվը տատանվում է 6-ից մինչև 1000 բջջի մեջ՝ կախված բակտերիաների տեսակից (նկ. 7)։
Նկ.7.Բակտերիաների մեջ դրոշակի տեղակայման տարբերակներ.
1 - monotrich, 2 - lophotrich;
3 - ամֆիտրիկ; 4 - peritrich.
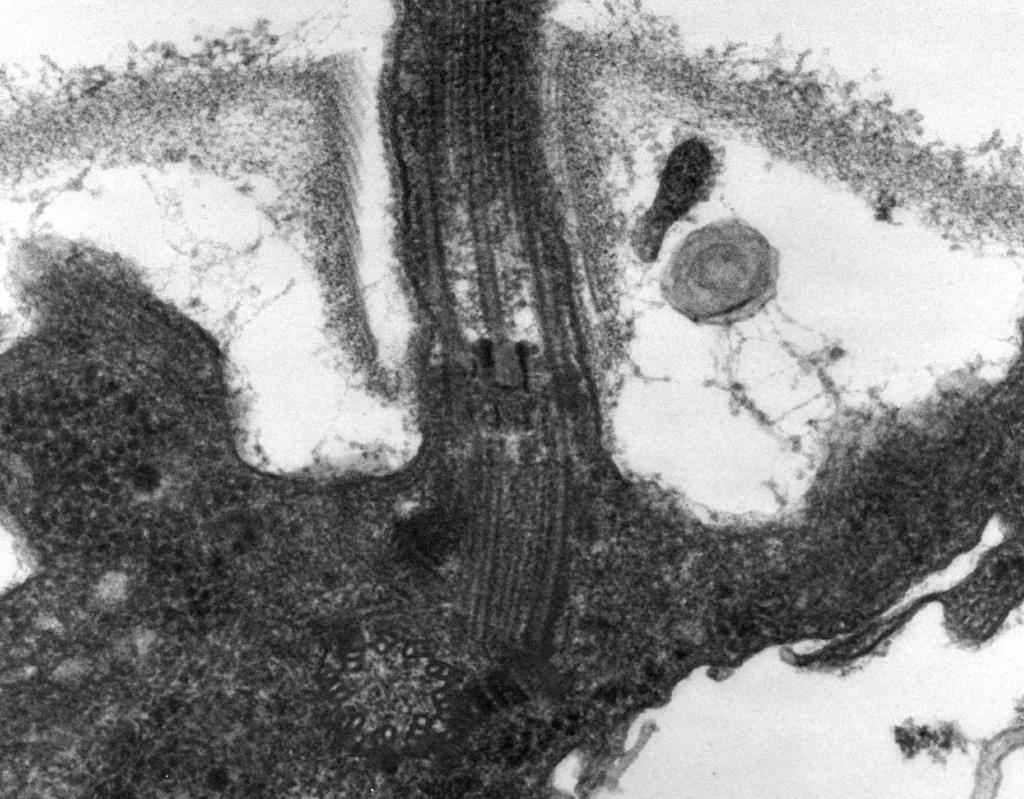
– Էլեկտրոնային մանրադիտակը ցույց է տալիս դա Դրոշակը բաղկացած է երեք մասից.պարուրաձև թել, կեռիկ և բազալ մարմին (նկ. 8):
Դրոշակի հիմնական մասը երկար է պարուրաձև թել (ֆիբրիլ) - մոտ 120 նմ տրամագծով կոշտ խոռոչ գլան, որը բաղկացած է flagellin սպիտակուցից: Սպիտակուցի մոլեկուլները թելի երկարությամբ կազմում են 11 շարք և դասավորված են պարուրաձև։ Թելի աճեցման ժամանակ բջջի ներսում սինթեզված սպիտակուցի մոլեկուլները անցնում են գլանակի խոռոչով և կցվում են պարույրի ծայրին։ Դրոշակի վերջում կա սպիտակուցի գլխարկ (կափարիչ), որը փակում է գլանի բացվածքը և կանխում սպիտակուցի մոլեկուլների արտազատումը շրջակա միջավայր: Դրոշակի թելի երկարությունը կարող է հասնել մի քանի միկրոմետրի: Բակտերիաների որոշ տեսակների մոտ դրոշակը լրացուցիչ ծածկված է արտաքինից պատյանով։ CS-ի մակերեսին պարուրաձև թելը անցնում է խտացած կոր կառուցվածքի՝ կեռիկի մեջ։

Բրինձ. ութ.Դրոշակի կառուցվածքի սխեման
2. Կեռիկ(20–45 նմ հաստությամբ) բջջի մակերեսին մոտ համեմատաբար կարճ գլան է, որը կազմված է ֆլագելլինից տարբերվող սպիտակուցից և ծառայում է թելքի ճկուն կապը բազալ մարմնի հետ։
3. Բազալային մարմինգտնվում է դրոշակի հիմքում և ապահովում է դրա պտույտը։ Բազալ մարմինը պարունակում է 9–12 տարբեր սպիտակուցներ և բաղկացած է երկու կամ չորս սկավառակներից (օղակներից)՝ ամրացված ձողի վրա, որը մանգաղի շարունակությունն է։ Այս օղակները տեղադրված են CPM-ում և CS-ում: Երկու ներքին օղակները (M և S) բազալ մարմնի պարտադիր բաղադրիչներն են։ M-օղակը տեղայնացված է CPM-ում, S-օղակը գտնվում է գրամ-բացասական բակտերիաների պերիպլազմիկ տարածության մեջ կամ գրամ-դրական բակտերիաների պեպտիդոգլիկան պարկի մեջ: Երկու արտաքին օղակները (D և L) ընտրովի են շարժման համար, քանի որ դրանք առկա են միայն գրամ-բացասական բակտերիաներում և տեղայնացված են, համապատասխանաբար, պեպտիդոգլիկան շերտում և CL-ի արտաքին թաղանթում: S, D և L օղակները անշարժ են և ծառայում են CS-ում դրոշակի ամրացմանը: Դրոշակի պտույտը որոշվում է բջջի CPM-ում ներկառուցված M օղակի պտույտով: Այսպիսով, դրոշակի բազալ մարմնի կառուցվածքային առանձնահատկությունները որոշվում են CS-ի կառուցվածքով:
Ֆունկցիոնալ առումով բազալ մարմինը էլեկտրական շարժիչ է, որն աշխատում է պրոտոններով: Բազալ մարմնի M օղակը (պտտվող ռոտոր) շրջապատված է բացասական լիցքերով թաղանթային սպիտակուցներով (շարժիչի ստատոր)։ Բակտերիալ բջիջն ունի էլեկտրաքիմիական էներգիան մեխանիկական էներգիայի փոխակերպելու արդյունավետ մեխանիզմ: Հետեւաբար, բակտերիան ծախսում է ամբողջ էներգիայի մոտ 0,1%-ը, որը ծախսում է դրոշակի աշխատանքի վրա։ Դրոշակի գործողության ժամանակ օգտագործվում է պրոտոն-շարժիչ ուժ, որն ապահովվում է մեմբրանի արտաքին և ներքին կողմերում պրոտոնների կոնցենտրացիաների տարբերությամբ (դրանք ավելի շատ են արտաքին կողմում) և ավելի բացասական լիցքի առկայությամբ։ թաղանթի ներքին կողմում: Պրոտոնի շարժիչ ուժը ստիպում է պրոտոններին բազալ մարմնի միջով անցնել բջիջ, մինչդեռ դրանք երկարաձգվում են ռոտորի որոշ մասերում՝ տալով նրանց դրական լիցք, այնուհետև պրոտոնները մտնում են բջիջ: Լիցքավորված հատվածները տեղակայված են այնպես, որ ռոտորի և ստատորի լիցքավորված հատվածների միջև առաջանում է գրավիչ ուժ, M-օղակը սկսում է պտտվել մոտ 300 պտ/րոպե արագությամբ։ Պտտման մեխանիզմ՝ COOH խմբի լիցքավորում-վերալիցքավորում ամինաթթուներում: Օղակի ամբողջական պտույտի համար բազալ մարմնով պետք է անցնի 500–1000 պրոտոն։ M-օղակի պտույտը դրա հետ կոշտ միացված առանցքի և կեռիկի միջով փոխանցվում է դրոշակի թելքին, որը գործում է որպես պտուտակ կամ նավի պտուտակ: Բակտերիան լողում է այնքան ժամանակ, քանի դեռ պտուտակն աշխատում է, իներցիայի ներդրումը չափազանց փոքր է։
Բացի այդ, բակտերիաները, նույնիսկ մեռածները, ջրային միջավայրում շարժվում են բրոունյան շարժման արդյունքում։ Բակտերիալ բջիջը մշտապես ենթարկվում է շրջակա մոլեկուլների ազդեցությանը, որոնք գտնվում են ջերմային շարժման մեջ: Տարբեր ուղղություններից հարվածները բակտերիային շպրտում են կողքից այն կողմ:
Դրոշակի շարժման տեսակը պտտվող է։ Շարժման երկու տեսակ կա՝ ուղղագիծ և պտտվող (շարժման ուղղության պարբերական պատահական փոփոխություններ)։ Երբ դրոշակները պտտվում են ժամացույցի սլաքի հակառակ ուղղությամբ (մոտ 1 վայրկյան), 40–60 ռ/րոպ հաճախականությամբ (միջին էլեկտրական շարժիչի արագությանը մոտ), դրանց թելերը հյուսվում են մեկ կապոցի մեջ (նկ. 9ա)։ Դրոշակի պտույտը փոխանցվում է բջիջին։ Քանի որ բջիջը շատ ավելի զանգված է, քան դրոշակը, այն սկսում է ուղիղ գծով շարժվել հակառակ ուղղությամբ՝ 3 անգամ պակաս արագությամբ, քան դրոշակի արագությունը։
Սա ապահովում է բջիջի թարգմանական շարժումը, որի արագությունը հեղուկ միջավայրում համար տարբեր տեսակներբակտերիաները 20–200 մկմ/վրկ են (սա համապատասխանում է րոպեում մարմնի մոտ 300–3000 երկարությանը) և ավելի դանդաղ շարժում պինդ միջավայրի մակերեսի վրա։
Բակտերիան կարող է նպատակաուղղված լողալ մեկ ուղղությամբ ոչ ավելի, քան 3 վրկ, այնուհետև շրջակա մոլեկուլների ազդեցությունը այն դարձնում է պատահական ուղղությամբ: Հետեւաբար, բակտերիաները մշակել են շարժման ուղղության ինքնաբուխ փոփոխության մեխանիզմ՝ դրոշակակիր շարժիչի միացում: Երբ այն սկսում է պտտվել ժամացույցի սլաքի ուղղությամբ (մոտ 0,1 վրկ), բակտերիան կանգ է առնում և շրջվում (կատարում է «թափել») պատահական ուղղությամբ: Այս դեպքում դրոշակները ցրվում են տարբեր ուղղություններով (նկ. 9բ): Ամֆիտրիկում շարժվելիս դրոշակի մեկ կապոցը շրջվում է ներսից դուրս (ինչպես քամուց ներս շրջված հովանոցը)։ Այնուհետև շարժիչը կրկին անցնում է ժամացույցի սլաքի ուղղությամբ ռոտացիայի, և բակտերիան նորից լողում է ուղիղ գծով, բայց այլ, պատահական ուղղությամբ:
Դրոշակը կարող է նաև փոխել ուղղությունը՝ ի պատասխան արտաքին գրգիռի: Եթե բակտերիան շարժվում է դեպի գրավիչի օպտիմալ կոնցենտրացիան, դրոշակները բջիջը մղում են միջավայրի միջով, նրա ուղղագիծ շարժումը դառնում է ավելի երկար, իսկ շրջվելու հաճախականությունը՝ ավելի ցածր, ինչը, ի վերջո, թույլ է տալիս նրան շարժվել ցանկալի ուղղությամբ:
Հայտնի են ոչ ակտիվ (կաթվածահար) դրոշակների առկայության դեպքեր։ Դրոշակավոր բակտերիաների շարժման համար անհրաժեշտ է ԿՍ-ի ամբողջականությունը (անխախտությունը): Բջիջների բուժումը լիզոզիմով, որը հանգեցնում է ԿՍ-ի պեպտիդոգլիկան շերտի հեռացմանը, հանգեցնում է բակտերիաների շարժվելու ունակության կորստի, թեև դրոշակները մնում են անձեռնմխելի:
Բակտերիաների տաքսիներ.Ցտեսություն միջավայրըմնում է անփոփոխ, բակտերիաները լողում են պատահական: Այնուամենայնիվ, շրջակա միջավայրը հազվադեպ է ամբողջովին միատարր: Եթե շրջակա միջավայրը տարասեռ է, բակտերիաները ցուցաբերում են տարրական վարքային ռեակցիաներ. նրանք ակտիվորեն շարժվում են որոշակի արտաքին գործոններով որոշված ուղղությամբ: Բակտերիաների գենետիկորեն որոշված նպատակային շարժումները կոչվում են տաքսիներ։ Կախված գործոնից՝ առանձնանում են քեմոտաքսիսը (հատուկ դեպք աերոտաքսիսն է), ֆոտոտաքսիսը, մագնետոտաքսիսը, թերմոտաքսիսը և վիսկոզիտաքսիսը։
Քիմոտաքսիս- շարժում որոշակի ուղղությամբ՝ կապված քիմիական նյութի աղբյուրի հետ. Քիմիական նյութերբաժանված են երկու խմբի՝ իներտ և պատճառող տաքսիներ՝ քիմոէֆեկտորներ։ Քիմիոէֆեկտորների շարքում կան բակտերիաներ գրավող նյութեր՝ գրավիչներ (շաքարներ, ամինաթթուներ, վիտամիններ, նուկլեոտիդներ), և դրանք վանող նյութեր՝ վանող նյութեր (որոշ ամինաթթուներ, սպիրտներ, ֆենոլներ, անօրգանական իոններ): Աերոբային և անաէրոբ պրոկարիոտների համար վանող նյութը մոլեկուլային թթվածինն է: Ներգրավիչները հաճախ ներկայացված են սննդային սուբստրատներով, թեև մարմնին անհրաժեշտ ոչ բոլոր նյութերն են գործում որպես գրավիչ: Բացի այդ, ոչ բոլոր թունավոր նյութերն են ծառայում որպես վանող և ոչ բոլոր վանողներն են վնասակար: Այսպիսով, բակտերիաները չեն կարողանում արձագանքել որևէ միացությունների, այլ միայն որոշակի և տարբեր բակտերիաների համար:
Բակտերիալ բջջի մակերևութային կառուցվածքներում կան հատուկ սպիտակուցային մոլեկուլներ՝ ընկալիչներ, որոնք հատուկ միանում են որոշակի քիմոէֆեկտորին, մինչդեռ քիմոէֆեկտորային մոլեկուլը չի փոխվում, բայց ընկալիչի մոլեկուլում տեղի են ունենում կոնֆորմացիոն փոփոխություններ: Ընկալիչները գտնվում են անհավասարաչափ բջջի ողջ մակերեսի վրա և կենտրոնացած են բևեռներից մեկում։ Ռեցեպտորի վիճակը արտացոլում է համապատասխան էֆեկտորի արտաբջջային կոնցենտրացիան:
Քեմոտաքսիսը հարմարվողական արժեք ունի: Օրինակ, խոլերայի վիբրիոյի ձևերը թույլ քիմոտաքսիսով ավելի քիչ վարակիչ են:
Աերոտաքսիս- Մոլեկուլային թթվածնի կարիք ունեցող բակտերիաները կուտակվում են օդային փուչիկների շուրջ, որոնք ընկել են ծածկույթի տակ:
Ֆոտոտաքսիս- շարժում դեպի լույս կամ հեռու լույս, որը բնորոշ է ֆոտոտրոֆ բակտերիաներին, որոնք օգտագործում են լույսը որպես էներգիայի աղբյուր:
մագնիտոտաքսիս- երկաթ պարունակող հանքանյութերի բյուրեղներ պարունակող ջրային բակտերիաների կարողությունը լողալու Երկրի մագնիսական դաշտի գծերով:
Թերմոտաքսիս- շարժումը ջերմաստիճանի փոփոխության ուղղությամբ, որն ունի մեծ նշանակությունորոշ պաթոգեն բակտերիաների համար.
Viscositaxis- լուծման մածուցիկության փոփոխություններին արձագանքելու ունակություն. Բակտերիաները սովորաբար հակված են դեպի ավելի բարձր մածուցիկություն ունեցող միջավայր, ինչը մեծ նշանակություն ունի պաթոգեն տեսակների համար:
Սահող բակտերիաներ.Փոքր արագությամբ (2–11 մկմ/վ) պինդ կամ մածուցիկ սուբստրատի վրայով սահելու ունակությունը հայտնաբերվել է որոշ պրոկարիոտների, օրինակ՝ միկոպլազմայի մոտ:
Կան մի քանի վարկածներ, որոնք բացատրում են սահող շարժումը։ Համաձայն ռեակտիվ շարժիչի վարկածդա պայմանավորված է լորձի արտազատմամբ բազմաթիվ լորձային ծակոտիների միջով SC-ում, որի արդյունքում բջիջը դուրս է մղվում ենթաշերտից լորձի արտազատման ուղղությամբ հակառակ ուղղությամբ: Համաձայն ճանապարհորդող ալիքի վարկածըՍահող շարժումը շարժական դրոշակներից ազատ ձևերով կապված է պեպտիդոգլիկան շերտի և CL-ի արտաքին թաղանթի միջև՝ պատվիրված մանրաթելերի բարակ սպիտակուցային շերտի առկայության հետ, որը նման է դրոշակի թելերին: Այս կառույցների կողմից «գործարկված» մանրաթելերի պտտվող շարժումը հանգեցնում է բջջի մակերևույթի վրա «շրջող ալիքի» (CL-ի շարժվող միկրոսկոպիկ ուռուցիկության) առաջացմանը, որի արդյունքում բջիջը վանվում է ենթաշերտից։ Վերջապես, որոշ սահող բակտերիաներում նկարագրված են կառուցվածքներ, որոնք նման են դրոշակավոր ձևերի բազալ մարմիններին:
Դրոշակների գործառույթները.
1. Ապահովել կպչունություն - սկզբնական փուլվարակիչ գործընթաց.
2. Ապահովել բակտերիաների շարժունակությունը։
3. Որոշեք հակագենային սպեցիֆիկությունը, սա H- հակագենն է:
Դրոշակների հայտնաբերում.
1. Մայրենի պատրաստուկների ֆազային կոնտրաստային մանրադիտակ («փշրված» և «կախված» կաթիլներ): Միկրոսկոպիկորեն շարժունակությունը որոշվում է ամենօրյա մշակույթի բջիջներում: Շարժունակությունը պասիվ բրոունյան շարժումից տարբերելու համար ուսումնասիրված մշակույթի մեկ կաթիլին ավելացվում է ֆենոլի 5% ջրային լուծույթի կաթիլ, այս դեպքում ակտիվ շարժումը դադարում է։
2. Մայրենի պատրաստուկների մութ դաշտային մանրադիտակ:
3. Ներկանյութերով կամ մետաղներով ներկված պատրաստուկների լուսային մանրադիտակ: Քանի որ պատրաստման ընթացքում դրոշակները շատ հեշտությամբ են վնասվում, այդ մեթոդները հազվադեպ են կիրառվում ամենօրյա պրակտիկայում:
Դրոշակները ներկվում են ագարի թեքության վրա աճեցված բջիջների միջոցով: Խտացման ջրի մոտ գտնվող բջիջները ընտրվում են բակտերիալ օղակով և զգուշորեն տեղափոխվում ստերիլ թորած ջրի մեջ, որը նույն ջերմաստիճանի է, ինչ բակտերիաների ինկուբացիայի ջերմաստիճանը թեք ագարի վրա, և բակտերիաները չեն թափահարվում օղակից, այլ զգուշորեն ընկղմվում են ջրի մեջ: Բակտերիաներով խողովակը թողնում են սենյակային ջերմաստիճանում 30 րոպե։ Օգտագործվում է քիմիապես մաքուր (քրոմի խառնուրդով լվացված) ապակի, որի վրա քսում են 2-3 կաթիլ կախոց։ Կախոցը տարածվում է ապակե մակերեսի վրա՝ նրբորեն թեքելով այն: Դեղը չորացրեք օդով:
Դրոշակները շատ բարակ են, ուստի դրանք կարող են հայտնաբերվել միայն հատուկ մշակման միջոցով: Սկզբում օֆորտի միջոցով ձեռք է բերվում այտուցվածություն և դրանց չափերի մեծացում, այնուհետև պատրաստումը գունավորվում է, ինչի շնորհիվ դրանք տեսանելի են դառնում լուսային մանրադիտակի տակ։
Ավելի հաճախ օգտագործվում է արծաթապատման մեթոդը ըստ Մորոզովի (նկ. 10).
- պատրաստուկը ամրացվում է սառցադաշտային քացախաթթվի լուծույթով 1 րոպե, լվանում ջրով;
- քսել տանինի լուծույթ (արևայրուք, դրոշակն ավելի խիտ դարձնելով) 1 րոպե, լվանալ ջրով;
- պատրաստումը մշակվում է 1-2 րոպե արծաթի նիտրատի ներծծող լուծույթով տաքացնելով, լվանում ջրով, չորացնում և մանրադիտակով մանրադիտակով:
Մանրադիտակով տեսանելի են մուգ շագանակագույն բջիջները և ավելի բաց դրոշակները:

Բրինձ. տասը.Դրոշակների նույնականացում արծաթապատմամբ
Բրինձ. տասնմեկ.Դրոշակների նույնականացում
էլեկտրոնային մանրադիտակով
Նկ. 4. Ծանր մետաղներով նստեցված պատրաստուկների էլեկտրոնային մանրադիտակ (նկ. 11):
5. Անուղղակի - ըստ բակտերիաների աճի բնույթի, երբ ցանվում է կիսահեղուկ 0,3% ագարում: 1-2 օր թերմոստատում մշակաբույսերի ինկուբացիայից հետո նշվում է բակտերիաների աճի բնույթը.
- ոչ շարժուն բակտերիաներում (օրինակ. S.saprophyticus) ներարկման երկայնքով աճ կա՝ «մեխ», իսկ միջավայրը թափանցիկ է.
շարժուն բակտերիաներում (օրինակ. E. coli) կա աճ ներարկման կողքին, ամբողջ ագարի սյունակում՝ «եղլնաձլ» և միջավայրի ցրված պղտորություն:
Բոլոր բակտերիաներըբաժանված է շարժական և անշարժ. Բակտերիաների շարժման օրգանները դրոշակ են։ Դրանք կազմված են ֆլագելինից՝ սպիտակուց, որը կառուցվածքային առումով կապված է միոզին տիպի կծկվող սպիտակուցների հետ։
Flagellum բազանբազալ մարմին է՝ բաղկացած սկավառակների համակարգից (բլեֆարոպլաստ՝ 1 սկավառակ՝ բջջային պատի արտաքին կողմը, 2 սկավառակ՝ բջջի պատի ներքին կողմը, 3 սկավառակ՝ ցիտոպլազմային թաղանթը), «ներկառուցված» ցիտոպլազմայում։ թաղանթ և բջջային պատ: Դրոշակի երկարությունը ավելի մեծ է, քան բուն միկրոբի մարմնի երկարությունը:
Ըստ դրոշակների քանակիև դրանց գտնվելու վայրը, շարժուն միկրոօրգանիզմները բաժանվում են.
1. Միաձույլ՝ մարմնի ծայրին մեկ դրոշակ ունեցող (առավել շարժուն)։ Օրինակ՝ Vibrio cholerae:
2. Լոֆոտրիխուս՝ բջջի բևեռներից մեկում դրոշակի կապոց: Օրինակ, Burkholderia (Pseudomonas) pseudomalei-ն մելիոիդոզի հարուցիչն է:
3. Ամֆիտրիկ՝ բջջի երկու բևեռներում դրոշակ ունեցող: Օրինակ՝ Spirillum volutans-ը։
4. Պերիտրիկ՝ բջջի ողջ պարագծի շուրջ դրոշակ ունեցող: Օրինակ, Escherichia coli, Salmonella typhi:
Դրոշակների հայտնաբերում.Դրոշակները շատ բարակ են, ուստի դրանք կարող են հայտնաբերվել միայն հատուկ մշակման միջոցով: Մասնավորապես, սկզբում մորդանտի միջոցով ձեռք է բերվում այտուցվածություն և դրանց չափերի մեծացում, ապա գունավորում են պատրաստուկը, ինչի շնորհիվ դրանք տեսանելի են դառնում լուսային մանրադիտակի տակ։ Դրոշակները կարելի է հայտնաբերել ըստ Մորոզովի, Լեֆլերի ներկման, ինչպես նաև էլեկտրոնային մանրադիտակի միջոցով։ Դրոշակները կարող են հայտնաբերվել նաև բակտերիաների ակտիվ շարժունակությամբ:
Մանրէների շարժումը դիտվում է կենդանի մշակույթներից «փշրված» և «կախված» կաթիլների պատրաստուկներում։ Այս պատրաստուկները մանրադիտակի են ենթարկվում չոր կամ ընկղմվող ոսպնյակով մութ դաշտում կամ ֆազային հակադրությամբ: Բացի այդ, շարժունակությունը կարող է որոշվել կիսահեղուկ ագարում բակտերիաների աճի բնույթով:
Խմել բակտերիայից.
Pili (pili), հոմանիշներ՝ villi, fimbria, սպիտակուցային բնույթի բարակ խոռոչ թելեր են, որոնք ծածկում են բակտերիաների բջիջների մակերեսը։ Ի տարբերություն դրոշակի, նրանք չեն կատարում շարժիչային ֆունկցիա։
Pili-ն տարածվում է բջջի մակերեսից և բաղկացած է սպիտակուցից պիլինա.
Ըստ իրենց գործառական նշանակության՝ դրանք բաժանվում են 2 տեսակի.
1) Առաջին տիպի կույտերը առկա են բակտերիաների մեծ մասում, ուստի դրանք կոչվում են «ընդհանուր տիպի վիլլի» (սովորական պիլին): Նրանք առաջացնում են բակտերիաների կցում կամ կպչում հյուրընկալող օրգանիզմի որոշակի բջիջներին։ Կպչունությունը ցանկացած վարակիչ գործընթացի սկզբնական փուլն է:
2) Երկրորդ տիպի պիլին (հոմանիշներ՝ կոնյուգատիվ, կամ սեռական՝ սեքս պիլի) հասանելի են միայն դոնոր բակտերիաներում, որոնք ունեն հատուկ պլազմիդ։ Նրանց թիվը փոքր է՝ 1-4 մեկ բջջում։
Հատակի սղոցները կատարում են հետևյալ գործառույթները.
1. Մասնակցել բակտերիաների միացման ժամանակ գենետիկական նյութի տեղափոխմանը մի բջջից մյուսը:
2. Նրանք կլանում են կոնկրետ բակտերիալ վիրուսներ՝ բակտերիոֆագներ
Բակտերիաների սպորները, ձևավորման պայմանները, Աուեսսկու ներկման գտնվելու վայրը, մեխանիզմը և փուլերը:
հակասություն- հանգստացող բակտերիաների յուրօրինակ ձև՝ բջջային պատի կառուցվածքի գրամ-դրական տեսակով:
սպորացում- սա արտաքին միջավայրում անբարենպաստ պայմաններում տեսակի (գենոֆորի) պահպանման միջոց է, այլ ոչ թե վերարտադրության մեթոդ։
Առաջանում են սպորներբակտերիաների գոյության համար անբարենպաստ պայմաններում (չորացում, սննդանյութերի անբավարարություն և այլն): Բակտերիալ բջջի ներսում ձևավորվում է մեկ սպոր (էնդոսպոր):
Սպորի ձևավորման փուլերը
1. Նախապատրաստական. Բակտերիաների ցիտոպլազմայում առաջանում է խտացված տարածք, որը չունի ազատ ջուր, որը կոչվում է «սպորոգեն գոտի», որը պարունակում է նուկլեոիդ։
2. Պրեսպորների (պրոսպորների) փուլ. Սպորածին գոտու շուրջ ձևավորվում է կրկնակի ցիտոպլազմային թաղանթի պատյան։
3. Պեպտիդոգլիկանից և կալցիումի աղերի և լիպիդների բարձր պարունակությամբ արտաքին թաղանթից կազմված կեղևի ձևավորում։
4. Հասունացման փուլ. Արտաքին թաղանթի արտաքին մասում ձևավորվում է սպորի ծրար, որից հետո լուծվում է բջջի վեգետատիվ մասը՝ ազատելով սպորը։
Մանրէաբանության զարգացումը վերջին տասնամյակների ընթացքում բազմաթիվ բացահայտումներ է բերել։ Եվ դրանցից մեկը դրոշակավոր բակտերիաների շարժման առանձնահատկություններն են։ Այս հնագույն օրգանիզմների շարժիչների դիզայնը պարզվեց, որ շատ բարդ է և, ըստ նրանց աշխատանքի սկզբունքի, շատ է տարբերվում նախակենդանիների մեր ամենամոտ էուկարիոտ հարազատների դրոշակներից: Դրոշակավոր մանրէի շարժիչը ամենաթեժ վեճն է եղել կրեացիոնիստների և էվոլյուցիոնիստների միջև: Բակտերիաների, դրանց դրոշակակիր շարժիչների և շատ ավելին - այս հոդվածը:
Ընդհանուր կենսաբանություն
Սկզբից հիշենք, թե ինչպիսի օրգանիզմներ են դրանք և ինչ տեղ են զբաղեցնում մեր մոլորակի օրգանական աշխարհի համակարգում։ Բակտերիաների տիրույթը միավորում է հսկայական քանակությամբ միաբջիջ պրոկարիոտ (առանց ձևավորված միջուկի) օրգանիզմների։
Այս կենդանի բջիջները հայտնվել են կյանքի ասպարեզում գրեթե 4 միլիարդ տարի առաջ և եղել են մոլորակի առաջին վերաբնակիչները: Նրանք կարող են լինել շատ տարբեր ձևերի (կոկիներ, ձողեր, վիբրիոներ, սպիրոխետներ), բայց դրանցից շատերը դրոշակավոր են։
Որտե՞ղ են ապրում բակտերիաները: Ամենուր. Մոլորակի վրա ապրում է ավելի քան 5 × 10 30: Դրանցից մոտ 40 միլիոն կա 1 գրամ հողում, մեր օրգանիզմում ապրում է մինչև 39 տրիլիոն: Դրանք կարելի է գտնել Մարիանայի խրամատի հատակում, օվկիանոսների հատակին տաք «սև ծխողների» մեջ, Անտարկտիդայի սառույցներում, և ներկայումս ձեր ձեռքերում կա մինչև 10 միլիոն բակտերիա:
Արժեքն անհերքելի է
Չնայած դրանց մանրադիտակային չափերին (0,5-5 մկմ), նրանց ընդհանուր կենսազանգվածը Երկրի վրա ավելի մեծ է, քան կենդանիների և բույսերի կենսազանգվածը միասին վերցրած: Նրանց դերը նյութերի շրջանառության մեջ անփոխարինելի է, իսկ սպառողների (օրգանական նյութերը ոչնչացնողների) հատկությունները թույլ չեն տալիս մոլորակը ծածկել դիակների լեռներով։
Դե, մի մոռացեք պաթոգենների մասին. ժանտախտի, ջրծաղիկի, սիֆիլիսի, տուբերկուլյոզի և շատ այլ վարակիչ հիվանդությունների հարուցիչները նույնպես բակտերիաներ են։
Բակտերիաները կիրառություն են գտել մարդու տնտեսական գործունեության մեջ։ Սկսած սննդի արդյունաբերությունից ( կաթնամթերքպանիրներ, թթու բանջարեղեն, ալկոհոլային խմիչքներ), «կանաչ» տնտեսությունը (կենսավառելիք և կենսագազ) դեպի բջիջների ճարտարագիտության և դեղերի արտադրության մեթոդներ (պատվաստանյութեր, շիճուկներ, հորմոններ, վիտամիններ):

Ընդհանուր մորֆոլոգիա
Ինչպես արդեն նշվեց, կյանքի այս միաբջիջ ներկայացուցիչները միջուկ չունեն, նրանց ժառանգական նյութը (ԴՆԹ-ի մոլեկուլները օղակի տեսքով) գտնվում են ցիտոպլազմայի (նուկլեոիդի) որոշակի տարածքում: Նրանց բջիջն ունի պլազմային թաղանթ և խիտ պարկուճ, որը ձևավորվում է պեպտիդոգլիկան մուրեյնի կողմից։ Բջջային օրգանելներից բակտերիաներն ունեն միտոքոնդրիաներ, կարող են լինել քլորոպլաստներ և տարբեր գործառույթներ ունեցող այլ կառուցվածքներ։
Բակտերիաների մեծ մասը դրոշակ է: Բջջի մակերևույթի ամուր պարկուճը թույլ չի տալիս նրանց տեղաշարժվել՝ փոխելով բջիջը, ինչպես անում են ամեոբաները: Նրանց դրոշակները տարբեր երկարությունների և մոտ 20 նմ տրամագծով խիտ սպիտակուցային գոյացություններ են։ Որոշ բակտերիաներ ունեն մեկ դրոշակ (միաձույլ), իսկ մյուսները ունեն երկու (ամֆիտրիկ): Երբեմն դրոշակները դասավորված են կապոցներով (լոֆոտրիխային) կամ ծածկում են բջջի ամբողջ մակերեսը (պերիտրիկ)։
Նրանցից շատերը ապրում են որպես միայնակ բջիջներ, բայց ոմանք կազմում են կլաստերներ (զույգեր, շղթաներ, թելեր, հիֆեր):

Շարժման առանձնահատկությունները
Դրոշակավոր բակտերիաները կարող են տարբեր կերպ շարժվել: Ոմանք շարժվում են միայն առաջ, իսկ ուղղությունը փոխում են շրջվելով: Ոմանք ունակ են կծկվել, մյուսները շարժվում են սահելով:
Բակտերիաների դրոշակները կատարում են ոչ միայն բջջային «թիակի» գործառույթներ, այլև կարող են լինել «բռնցքամարտի» գործիք։
Մինչև վերջերս ենթադրվում էր, որ բակտերիաների դրոշակը շարժվում է օձի պոչի պես: Վերջին ուսումնասիրությունները ցույց են տվել, որ բակտերիաների դրոշակը շատ ավելի բարդ է: Այն աշխատում է տուրբինի պես։ Կցված շարժիչին, այն պտտվում է մեկ ուղղությամբ: Գործարկիչը կամ բակտերիաների դրոշակակիր շարժիչը բարդ մոլեկուլային կառուցվածք է, որն աշխատում է մկանի նման: Այն տարբերությամբ, որ կծկվելուց հետո մկանը պետք է թուլանա, իսկ բակտերիալ շարժիչն անընդհատ աշխատում է։

Դրոշակի նանոմեխանիզմը
Չխորանալով շարժման կենսաքիմիայի մեջ՝ մենք նշում ենք, որ դրոշակի շարժման ստեղծմանը մասնակցում են մինչև 240 սպիտակուցներ, որոնք բաժանված են 50 մոլեկուլային բաղադրիչների՝ համակարգում հատուկ գործառույթով:
Բակտերիաների այս շարժիչ համակարգում կա ռոտոր, որը շարժվում է, և ստատոր, որն ապահովում է այդ շարժումը: Առկա է շարժիչ լիսեռ, բուշ, կցորդիչ, արգելակներ և արագացուցիչներ
Այս մանրանկարիչ շարժիչը թույլ է տալիս բակտերիաներին 35 անգամ գերազանցել իրենց չափը ընդամենը 1 վայրկյանում: Միևնույն ժամանակ, բուն դրոշակի աշխատանքը, որը րոպեում կատարում է 60 հազար պտույտ, օրգանիզմը ծախսում է բջիջի սպառած ողջ էներգիայի միայն 0,1%-ը։
Զարմանալի է նաև, որ բակտերիան կարող է փոխարինել և վերանորոգել իր շարժիչի մեխանիզմի բոլոր պահեստամասերը «երթևեկելիս»: Պարզապես պատկերացրեք, որ դուք ինքնաթիռում եք: Իսկ տեխնիկները փոխում են աշխատող շարժիչի սայրերը:
Flagellate բակտերիան ընդդեմ Դարվինի
Շարժիչը, որը կարող է աշխատել մինչև 60,000 պտույտ/րոպե արագությամբ, ինքնագործարկվել և որպես վառելիք օգտագործել միայն ածխաջրեր (շաքար), ունենալով էլեկտրական շարժիչի նման սարք, կարո՞ղ է նման սարքը առաջանալ էվոլյուցիայի գործընթացում:
Սա այն հարցն է, որ ինքն իրեն տվել է կենսաբանության գիտությունների թեկնածու Մայքլ Բեհեն 1988թ. Նա կենսաբանության մեջ մտցրեց անկրճատելի համակարգի հայեցակարգը. մի համակարգ, որի բոլոր մասերը միաժամանակ անհրաժեշտ են դրա գործունեությունը ապահովելու համար, և առնվազն մեկ մասի հեռացումը հանգեցնում է նրա գործունեության ամբողջական խաթարմանը:
Դարվինի էվոլյուցիայի տեսանկյունից մարմնի բոլոր կառուցվածքային փոփոխությունները տեղի են ունենում աստիճանաբար, և միայն հաջողակներն են ընտրվում բնական ընտրության միջոցով:
Մ. Բեհեի եզրակացությունները, որոնք ներկայացված են «Դարվինի սև արկղը» գրքում (1996 թ.). համակարգ, ինչը նշանակում է, որ այս համակարգը չէր կարող առաջանալ բնական ընտրության միջոցով:

Բալզամ կրեացիոնիստների համար
Ստեղծագործության տեսությունը, ինչպես ներկայացրել է գիտնական և կենսաբանության պրոֆեսոր, Բեթղեհեմի (ԱՄՆ) Լիհայ համալսարանի կենսաբանական գիտությունների ֆակուլտետի դեկան Մ. Բեհեն, անմիջապես գրավեց եկեղեցու սպասավորների և աստվածային տեսության կողմնակիցների ուշադրությունը։ կյանքի ծագումը.
2005-ին Միացյալ Նահանգներում նույնիսկ դատական հայց կար, որտեղ Բեհեն վկա էր «ողջամիտ դիզայնի» տեսության կողմնակիցներից, որը դիտարկում էր կրեացիոնիզմի ուսումնասիրության ներդրումը Դովերի դպրոցներում դասընթացում « Պանդաների և մարդկանց վրա»: Գործընթացը կորավ, նման առարկայի դասավանդումը ճանաչվեց գործող սահմանադրությանը հակասող։
Սակայն կրեացիոնիստների և էվոլյուցիոնիստների միջև բանավեճը շարունակվում է այսօր:
Դրոշակը բակտերիաների բջիջի մակերեսային կառուցվածքն է, որը ծառայում է նրանց հեղուկ միջավայրում տեղաշարժվելու համար:
Կախված դրոշակի տեղակայությունից՝ բակտերիաները բաժանվում են (նկ. 1).
Պերիտրիխիալ
խառը
բեւեռ
Ենթաբևեռ
Բևեռային դրոշակ- մեկ կամ մի քանի դրոշակ գտնվում են բջջի մեկ (միաբևեռ) կամ երկու (երկբևեռ) բևեռների վրա, և հիմքը զուգահեռ է բջջի երկար առանցքին:
Ենթաբևեռ դրոշակ(ենթաբևեռ) - մեկ կամ մի քանի դրոշակ գտնվում է կողային մակերեսի անցման կետում բջջի մեկ կամ երկու ծայրերում: Հիմքում ուղիղ անկյուն է բջջի երկար առանցքի հետ:
Կողային դրոշակ(կողային) - մեկ կամ մի քանի դրոշակ կապոցի տեսքով գտնվում են բջջի կեսերից մեկի միջնակետում:
Peritrichial flagella- գտնվում է բջջի ամբողջ մակերեսի վրա մեկ առ մեկ կամ կապոցներով, բևեռները սովորաբար զրկված են դրանցից:
Խառը դրոշակ- երկու կամ ավելի դրոշակներ են գտնվում տարբեր կետերբջիջները.
Կախված դրոշակների քանակից՝ առանձնանում են.
Միաձույլ - մեկ դրոշակ
Polytrichs - դրոշակների փունջ
Նաև առանձնանում են.
լոֆոտրիկ- Դրոշակների մենաշնորհային պոլիտրիխային դասավորություն.
ամֆիտրիխներ- Դրոշակների երկբևեռ պոլիտրիխիալ դասավորություն.
Բակտերիալ դրոշակի և բազալ մարմնի կառուցվածքը: Flagellum.
Դրոշակն ինքնին դասավորված է բավականին պարզ՝ մի թել, որը կցված է բազալ մարմնին: Երբեմն խողովակի կոր հատվածը, այսպես կոչված, կեռիկը կարող է տեղադրվել բազալ մարմնի և թելքի միջև, այն ավելի հաստ է, քան թելիկը և մասնակցում է թելքի ճկուն ամրացմանը բազալ մարմնին:
Քիմիական բաղադրության առումով դրոշակը բաղկացած է 98% ֆլագելինի սպիտակուցից (flagellum - flagellum), պարունակում է 16 ամինաթթուներ, գերակշռում են գլուտամին և ասպարտիկ թթուները, տրիպտոֆանը, ցիստեինը և ցիստինը բացակայում են անուշաբույր ամինաթթուների փոքր քանակությամբ: Flagellin-ը պարտադրում է հակագենային առանձնահատկություն, այն կոչվում է H-հակագին: Բակտերիալ դրոշակները չունեն ATPase ակտիվություն:
Դրոշակի հաստությունը 10–12 նմ է, երկարությունը՝ 3–15 մկմ։
Այն կոշտ պարույր է՝ ժամացույցի սլաքի ուղղությամբ ոլորված։ Դրոշակի պտույտը կատարվում է նաև ժամացույցի սլաքի ուղղությամբ՝ 40 պտ/րոպից մինչև 60 պտ/րոպ հաճախականությամբ, ինչը հանգեցնում է բջիջի պտտման հակառակ ուղղությամբ, բայց քանի որ բջիջը շատ ավելի ծանր է, քան դրոշակը, այնուհետև դրա պտույտը դանդաղ է 12-ից մինչև 14 ռ / րոպե:
Դրոշակը աճում է հեռավոր ծայրից, որտեղ ստորաբաժանումները մտնում են ներքին ալիքով։ Որոշ տեսակների մոտ դրոշակը արտաքինից լրացուցիչ ծածկված է պատյանով, որը բջջային պատի շարունակությունն է և հավանաբար ունի նույն կառուցվածքը։
Բազալային մարմին
Բազալ մարմինը բաղկացած է 4 մասից.
Ձողերի զուգավորում թելիկով կամ կեռիկով
Գավազանի վրա ամրացված երկու սկավառակ: (M և S)
Սպիտակուցային համալիրների խումբ (ստատորներ)
սպիտակուցային գլխարկ
Բակտերիաները, որոնք ունեն ներքին և արտաքին թաղանթ, ունեն 2 լրացուցիչ սկավառակներ (P և L) և սպիտակուցային կառուցվածքներ, որոնք գտնվում են արտաքին թաղանթի վրա՝ բազալ մարմնի մոտ, հետևաբար նրանք կարևոր դեր չեն խաղում շարժման մեջ։
Բազալային մարմնի կառուցվածքի առանձնահատկությունը որոշվում է բջջային պատի կառուցվածքով. դրա անձեռնմխելիությունը անհրաժեշտ է դրոշակի շարժման համար: Բջիջների լիզոզիմով բուժումը հանգեցնում է պեպտիդոգլիկանի շերտի հեռացմանը բջջային պատից, ինչը հանգեցնում է շարժման կորստի, թեև դրոշակի կառուցվածքը չի խախտվել։
Բակտերիալ դրոշակները որոշում են բակտերիալ բջջի շարժունակությունը: Դրոշակները բարակ թելեր են, որոնք առաջանում են ցիտոպլազմային թաղանթից և ավելի երկար են, քան բջջը: Դրոշակները ունեն 12–20 նմ հաստություն և 3–15 մկմ երկարություն։ Դրանք բաղկացած են 3 մասից՝ պարուրաձև թելից, կեռիկից և հատուկ սկավառակներով ձող պարունակող բազալ մարմնից (1 զույգ սկավառակ գրամ դրական և 2 զույգ գրամ-բացասական բակտերիաների համար)։ Դրոշակի սկավառակները կցվում են ցիտոպլազմային թաղանթին և բջջային պատին։ Սա ստեղծում է էլեկտրական շարժիչի ազդեցությունը շարժիչի գավազանով, որը պտտում է դրոշակը: Դրոշակները բաղկացած են սպիտակուցից՝ flagellin-ից (flagelum - flagellum); H անտիգեն է: Ֆլագելինի ենթամիավորները ոլորված են: Դրոշակների թիվը տարբեր տեսակների բակտերիաներում տատանվում է մեկից (միաձույլ) Vibrio cholerae-ում մինչև տասը կամ հարյուրավոր դրոշակներ, որոնք տարածվում են բակտերիաների պարագծի երկայնքով Escherichia coli-ում, Proteus-ում և այլն: խցիկի վերջը. Ամֆիտրիկները բջջի հակառակ ծայրերում ունեն մեկ դրոշակ կամ դրոշակի փաթեթ:
Pili (fimbriae, villi) - թելավոր գոյացություններ, ավելի բարակ և կարճ (3-10 նմ x 0,3-10 մկմ), քան դրոշակները: Pili-ն տարածվում է բջջային մակերեսից և բաղկացած է պիլին սպիտակուցից, որն ունի հակագենային ակտիվություն: Կան կպչունություն, այսինքն՝ ախտահարված բջիջին բակտերիաներ կցելու համար, ինչպես նաև սնուցման, ջրային աղի նյութափոխանակության և սեռական (F-pili) կամ խոնարհման համար պատասխանատու պիլիներ։ Խմիչքները շատ են՝ մեկ վանդակի համար մի քանի հարյուր: Այնուամենայնիվ, սեռի սեռը սովորաբար 1-3 է մեկ բջջում. դրանք ձևավորվում են այսպես կոչված «արական» դոնոր բջիջներով, որոնք պարունակում են փոխանցվող պլազմիդներ (F-, R-, Col-պլազմիդներ): Սեռական պիլի տարբերակիչ առանձնահատկությունը փոխազդեցությունն է հատուկ «արական» գնդաձև բակտերիոֆագների հետ, որոնք ինտենսիվորեն ներծծվում են սեքս պիլի վրա:
Սպորները քնած ամուր բակտերիաների յուրահատուկ ձև են, այսինքն. գրամ դրական բջջային պատի կառուցվածքով բակտերիաներ. Սպորները ձևավորվում են բակտերիաների գոյության համար անբարենպաստ պայմաններում (չորացում, սննդանյութերի պակաս և այլն: Բակտերիաների բջջի ներսում ձևավորվում է մեկ սպոր (էնդոսպոր): Սպորների առաջացումը նպաստում է տեսակների պահպանմանը և վերարտադրության մեթոդ չէ: , ինչպես սնկերում: Bacillus սեռի սպոր առաջացնող բակտերիաները ունեն սպորներ, որոնք չեն գերազանցում բջջի տրամագիծը: Բակտերիաները, որոնց սպորի չափը գերազանցում է բջջի տրամագիծը, կոչվում են կլոստրիդիաներ, օրինակ՝ Clostridium սեռի բակտերիաները (լատ. Clostridium - spindle): Սպորները թթու դիմացկուն են, հետևաբար, դրանք կարմիր ներկվում են ըստ Aujeszky մեթոդի կամ ըստ Ziehl-Neelsen մեթոդի, իսկ վեգետատիվ բջիջը կապույտ:
Վեճի ձևը կարող է լինել օվալաձև, գնդաձև; խցում գտնվելու վայրը տերմինալ է, այսինքն. փայտիկի վերջում (տետանուսի հարուցիչում), ենթատերմինալ - ավելի մոտ փայտի ծայրին (բոտուլիզմի, գազային գանգրենա ախտածինների մոտ) և կենտրոնական (սիբիրախտի բացիլներում): Սպորը երկար ժամանակ պահպանվում է բազմաշերտ կեղևի, կալցիումի դիպիկոլինատի, ցածր ջրի պարունակության և նյութափոխանակության դանդաղ պրոցեսների առկայության պատճառով: Բարենպաստ պայմաններում սպորները բողբոջում են երեք հաջորդական փուլերով՝ ակտիվացում, սկիզբ, բողբոջում։
8. Բակտերիաների հիմնական ձեւերը
Գնդիկավոր բակտերիաներ (կոկիներ)սովորաբար գնդաձև են, բայց կարող են լինել թեթևակի օվալաձև կամ լոբի տեսքով: Cocci-ն կարող է տեղակայվել առանձին (micrococci); զույգերով (diplococci); շղթաների (streptococci) կամ խաղողի ողկույզների (staphylococci) տեսքով, փաթեթ (sarcinas): Streptococci-ն կարող է առաջացնել տոնզիլիտ և erysipelas, staphylococci՝ տարբեր բորբոքային և թարախային պրոցեսներ։
ձողաձև բակտերիաներառավել տարածված. Ձողերը կարող են լինել միայնակ, միացված զույգերով (դիպլոբակտերիա) կամ շղթայական (streptobacteria): Ձողաձև բակտերիաները ներառում են Escherichia coli-ն, սալմոնելոզի, դիզենտերիային, որովայնային տիֆի, տուբերկուլյոզի և այլնի հարուցիչները: Որոշ ձողաձև բակտերիաներ ունեն անբարենպաստ պայմաններում ձևավորվելու հատկություն: վեճեր.Սպոր առաջացնող ձողերը կոչվում են բացիլներ.Սուրաձև բացիլները կոչվում են կլոստրիդիա.
Սպորացումը բարդ գործընթաց է: Սպորները զգալիորեն տարբերվում են սովորական բակտերիալ բջջից: Նրանք ունեն խիտ թաղանթ և շատ քիչ քանակությամբ ջուր, սննդարար նյութեր չեն պահանջում, և վերարտադրությունը լիովին դադարում է։ Սպորները երկար ժամանակ կարող են դիմակայել չորացմանը, բարձր և ցածր ջերմաստիճաններին և կարող են կենսունակ վիճակում լինել տասնյակ և հարյուրավոր տարիներ (սիբիրախտի սպորներ, բոտուլիզմ, տետանուս և այլն): Բարենպաստ միջավայրում հայտնվելով՝ սպորները բողբոջում են, այսինքն՝ վերածվում են սովորական վեգետատիվ բազմացման ձևի։
Խճճված բակտերիաներկարող է լինել ստորակետի տեսքով՝ վիբրիոներ, մի քանի գանգուրներով՝ սպիրիլլա, բարակ ոլորված փայտիկի տեսքով՝ սպիրոխետներ։ Վիբրիոնները խոլերայի հարուցիչն են, իսկ սիֆիլիսի հարուցիչը սպիրոխետն է։
9. Ռիկեցիայի և քլամիդիայի մորֆոլոգիայի առանձնահատկությունները
Ռիկեցիաները փոքր գրամ-բացասական միկրոօրգանիզմներ են, որոնք բնութագրվում են ընդգծված պոլիմորֆիզմով. ձևավորում են կոկիկներ, ձողաձև և թելիկ ձևեր (նկ. 22): Ռիկեցիայի չափերը տատանվում են 0,5-ից մինչև 3-4 մկմ, թելային ձևերի երկարությունը հասնում է 10-40 մկմ-ի։ Նրանք սպորներ և պարկուճներ չեն առաջացնում, կարմիր ներկված են ըստ Զդրոդովսկու։
Քլամիդիաները գնդաձեւ են, ձվաձեւ կամ ձողաձեւ։ Նրանց չափերը տատանվում են 0,2-1,5 մկմ սահմաններում։ Քլամիդիայի մորֆոլոգիան և չափերը կախված են դրանց ներբջջային զարգացման ցիկլի փուլից, որը բնութագրվում է փոքր գնդաձև տարրական գոյացության վերափոխմամբ երկուական բաժանմամբ մեծ սկզբնական մարմնի։ Նախքան բաժանումը, քլամիդիայի մասնիկները պարուրված են բակտերիալ պարկուճ հիշեցնող գոյացության մեջ: Chlamydia բիծը «* ըստ Ռոմանովսկի-Գիմսայի, գրամ-բացասական, հստակ տեսանելի է ինտրավիտալ պատրաստուկներում ֆազային կոնտրաստային մանրադիտակով:
10. Միկոպլազմայի կառուցվածքը և կենսաբանությունը:
Միայն մեկ կարգ՝ Mycoplasmatales, պատկանում է Mollicutes դասին։ Այս կարգի ներկայացուցիչները՝ միկոպլազմաները.
Նրանք բակտերիայից տարբերվում են նրանով, որ չունեն բջջային պատ։ Փոխարենը դրանք պարունակում են եռաշերտ լիպոպրոտեինային ցիտոպլազմային թաղանթ: Միկոպլազմայի չափերը տատանվում են 125-250 մկմ-ի սահմաններում։ Կլոր են, օվալաձև կամ թելիկ, գրամ-բացասական։
Միկոպլազմաները բազմանում են երկուական տրոհմամբ, ինչպես բակտերիաների մեծ մասը, հատկապես թելիկ կառուցվածքներում փոքր կոկոիդային գոյացությունների (տարրական մարմիններ, EB) ձևավորումից հետո։
Միկոպլազմաները ունակ են բողբոջելու և բաժանվելու: Նվազագույն վերարտադրող միավորը ET է (0,7-0,2 մկմ): Բջջային թաղանթի հիմնական բաղադրիչը խոլեստերինն է։ Միկոպլազմաները ունակ չեն խոլեստերին ձևավորելու և այն օգտագործում են հյուսվածքներից կամ սննդանյութերից, որոնք լրացվում են դրանց ներմուծմամբ: Գրամ ներկումը բացասական է, բայց Ռոմանովսկի-Գիմսա ներկումը տալիս է լավագույն արդյունքը: Միկոպլազմաները պահանջում են աճեցման պայմաններ. բնական շիճուկ, խոլեստերին, նուկլեինաթթուներ, ածխաջրեր, վիտամիններ և տարբեր աղեր պետք է ավելացվեն սննդարար միջավայրում: Խիտ միջավայրերի վրա նրանք ձևավորում են բնորոշ փոքր կիսաթափանցիկ գաղութներ՝ բարձրացված հատիկավոր կենտրոնով՝ տալով նրանց «տապակած ձվերի» տեսք։ Արյունով միջավայրում միկոպլազմայի որոշ տեսակներ տալիս են ա- և բետա-հեմոլիզ: Կիսահեղուկ միջավայրում միկոպլազմաները աճում են ներարկման գծի երկայնքով՝ ձևավորելով ցրված, փխրուն գաղութներ։ Հեղուկ միջավայրում դրանք հանգեցնում են թեթև մառախուղի կամ բծախնդրության; որոշ շտամներ կարողանում են ձևավորել ամենաբարակ յուղոտ թաղանթը: Մարդկանց մեջ առանձնացված են Mycoplasma, Ureaplasma և Acholeplasma սեռերի ներկայացուցիչները, ներառյալ պաթոգեն և սապրոֆիտ տեսակները: