ตามตำแหน่งของแฟลกเจลลา แบคทีเรียจะถูกแบ่งออกเป็น Organelles ของการเคลื่อนไหว: หน้าที่และโครงสร้างคุณสมบัติของการเคลื่อนไหวของโปรโตซัว
โครงสร้าง. ประมาณครึ่งหนึ่งของแบคทีเรียที่รู้จักบนพื้นผิวมีอวัยวะของการเคลื่อนไหว - แฟลกเจลลาโค้งคล้ายคลื่น มวลของแฟลกเจลลานั้นสูงถึง 2% ของมวลแห้งของแบคทีเรีย ความยาวของแฟลเจลลัมนั้นยาวกว่าความยาวของลำตัวของจุลินทรีย์และเท่ากับ 3–12 ไมโครเมตร ความหนาของแฟลเจลลัมคือ 0.02 ไมโครเมตร และแฟลเจลลามีขั้วจะหนากว่าแฟลเจลลัม
แฟลกเจลลาประกอบด้วยโปรตีนแฟลเจลลิน (ลาตินแฟลเจลลา - แฟลเจลลัม) ซึ่งในโครงสร้างของมันเป็นโปรตีนหดตัวประเภทไมโอซิน แฟลเจลลัมประกอบด้วยเส้นใยโปรตีนที่เป็นเนื้อเดียวกันหนึ่งเส้นหรือเส้นใย 2-3 เส้นที่พับเป็นเปียอย่างแน่นหนา ด้ายของแฟลเจลลัมเป็นเกลียวแข็งบิดทวนเข็มนาฬิกา ระยะห่างของเกลียวนั้นจำเพาะสำหรับแบคทีเรียแต่ละชนิด
จำนวน ขนาด และการจัดเรียงของแฟลกเจลลาเป็นคุณลักษณะที่คงที่สำหรับสปีชีส์หนึ่งๆ และนำมาพิจารณาในอนุกรมวิธาน อย่างไรก็ตาม แบคทีเรียบางชนิดสามารถสร้างแฟลกเจลลาได้ ประเภทต่างๆ. นอกจากนี้ การปรากฏตัวของแฟลกเจลลาขึ้นอยู่กับสภาวะแวดล้อม: บนสื่อที่เป็นของแข็งในระหว่างการเพาะเลี้ยงระยะยาว แบคทีเรียสามารถสูญเสียแฟลกเจลลา และในสื่อที่เป็นของเหลวก็สามารถได้รับอีกครั้ง จำนวนและตำแหน่งของแฟลกเจลลาในสายพันธุ์เดียวกันสามารถกำหนดได้โดยระยะของวงจรชีวิต ดังนั้น ความสำคัญทางอนุกรมวิธานของคุณลักษณะนี้ไม่ควรประเมินค่าสูงไป
การจำแนกแบคทีเรียตามจำนวนและตำแหน่งของแฟลกเจลลา:
1. อาทริชิ - แฟลกเจลลาไม่อยู่
2. Monotricus- แฟลเจลลัมหนึ่งตัวอยู่ที่เสาหนึ่งเซลล์ (สกุล วิบริโอ)- การจัดเรียง monopolar monotrichous ของ flagella ซึ่งเป็นแบคทีเรียที่เคลื่อนที่ได้มากที่สุด
3. Polytricus - แฟลกเจลลามากมาย:
– lophorichous- มัดแฟลกเจลลาที่ขั้วหนึ่งของเซลล์ (การเกิด ซูโดโมนาส, Burkholderia) - การจัดเรียง monopolar polytrichous ของ flagella;
– สะเทินน้ำสะเทินบก- ที่แต่ละขั้วของเซลล์มีมัดแฟลเจลลัม (สกุล สไปริลลัม)- การจัดเรียงตัวของแฟลเจลลาสองขั้ว polytrichous;
– peririchous- แฟลกเจลลาถูกจัดเรียงอย่างไม่มีลำดับเฉพาะบนพื้นผิวทั้งหมดของเซลล์ (fam. Enterobacteriaceae(การเกิด Escherichia, โพรทูส) แฟม. แบคทีเรีย,ตระกูล Clostidiaceae) จำนวนแฟลกเจลลาแตกต่างกันไปตั้งแต่ 6 ถึง 1,000 ต่อเซลล์ ขึ้นอยู่กับชนิดของแบคทีเรีย (รูปที่ 7)
รูปที่ 7ตัวเลือกสำหรับตำแหน่งของแฟลกเจลลาในแบคทีเรีย:
1 - monotrich, 2 - lophorich;
3 - สะเทินน้ำสะเทินบก; 4 - เปริทริช
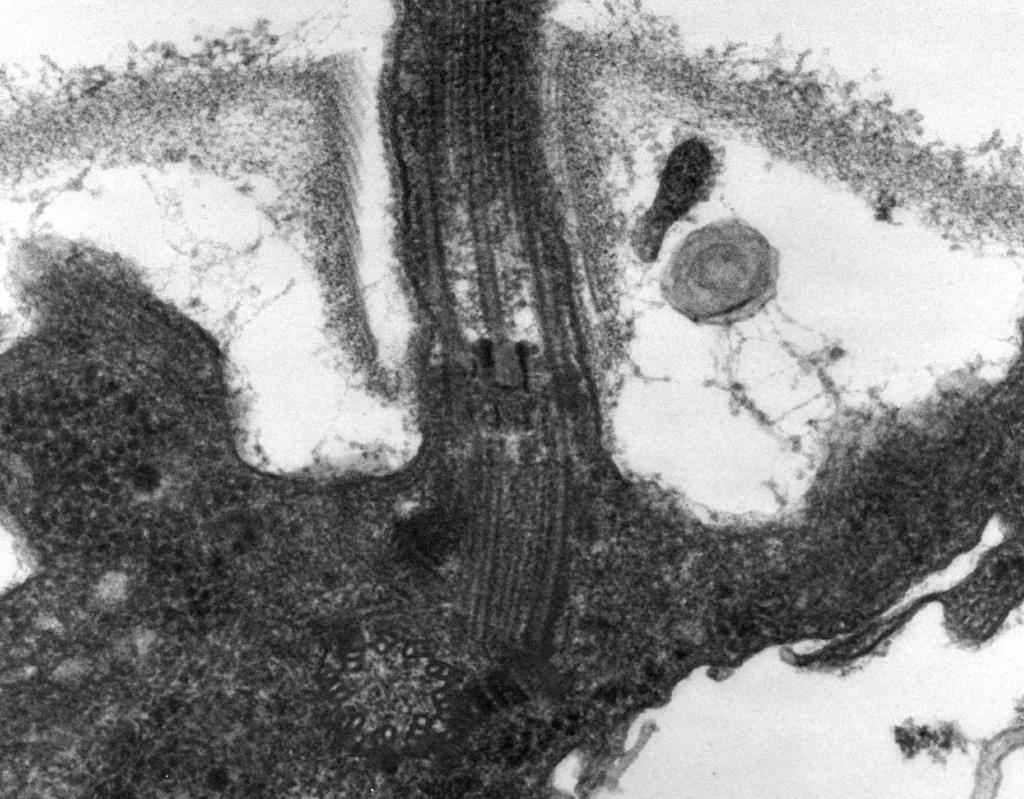
– กล้องจุลทรรศน์อิเล็กตรอนเผยว่า แฟลเจลลัมประกอบด้วยสามส่วน:ไส้เกลียว ตะขอ และตัวฐาน (รูปที่ 8)
ส่วนหลักของแฟลเจลลัมนั้นยาว เกลียวเกลียว (ไฟบริล) - ทรงกระบอกกลวงแข็งที่มีขนาดเส้นผ่าศูนย์กลางประมาณ 120 นาโนเมตรประกอบด้วยโปรตีนแฟลเจลลิน โมเลกุลของโปรตีนก่อตัวเป็น 11 แถวตามความยาวของเกลียวและจัดเรียงเป็นเกลียว ในระหว่างการเจริญเติบโตของเส้นด้าย โมเลกุลโปรตีนที่สังเคราะห์ขึ้นภายในเซลล์จะผ่านโพรงของกระบอกสูบและยึดติดกับเกลียวที่ปลาย ที่ส่วนท้ายของแฟลเจลลัมจะมีฝาครอบโปรตีน (ฝา) ที่ปิดช่องเปิดของกระบอกสูบและป้องกันการปล่อยโมเลกุลโปรตีนออกสู่สิ่งแวดล้อม ความยาวของไส้หลอดแฟลเจลลัมสามารถเข้าถึงได้หลายไมโครเมตร ในแบคทีเรียบางชนิด แฟลเจลลัมยังถูกหุ้มด้วยฝักด้านนอกอีกด้วย ที่พื้นผิวของ CS เกลียวเกลียวจะผ่านเข้าไปในโครงสร้างโค้งหนา - เบ็ด

ข้าว. แปด.แผนผังโครงสร้างของแฟลเจลลัม
2. ตะขอ(หนา 20–45 นาโนเมตร) ใกล้ผิวเซลล์เป็นทรงกระบอกที่ค่อนข้างสั้น ซึ่งประกอบด้วยโปรตีนที่แตกต่างจากแฟลเจลลิน และทำหน้าที่ให้การเชื่อมต่อที่ยืดหยุ่นของไส้หลอดกับตัวฐาน
3. ฐานร่างกายตั้งอยู่ที่ฐานของแฟลเจลลัมและทำให้แน่ใจว่าหมุนได้ ตัวฐานประกอบด้วยโปรตีนที่แตกต่างกัน 9–12 ชนิดและประกอบด้วยดิสก์ (วงแหวน) สองหรือสี่อันที่ร้อยอยู่บนไม้เรียว ซึ่งเป็นส่วนต่อของตะขอ วงแหวนเหล่านี้ติดตั้งอยู่ใน CPM และ CS วงแหวนด้านในสองวง (M และ S) เป็นส่วนประกอบบังคับของตัวฐาน M-ring ถูกแปลเป็นภาษาท้องถิ่นใน CPM, S-ring ตั้งอยู่ในปริภูมิ periplasmic ของแบคทีเรียแกรมลบหรือในถุง peptidoglycan ของแบคทีเรียแกรมบวก วงแหวนรอบนอกสองวง (D และ L) เป็นทางเลือกสำหรับการเคลื่อนไหว เนื่องจากมีอยู่ในแบคทีเรียแกรมลบเท่านั้นและถูกแปลเป็นภาษาท้องถิ่นตามลำดับ ในชั้น peptidoglycan และในเยื่อหุ้มชั้นนอกของ CL วงแหวน S, D และ L ไม่สามารถเคลื่อนที่ได้และทำหน้าที่ซ่อมแฟลเจลลัมใน CS การหมุนของแฟลเจลลัมถูกกำหนดโดยการหมุนของวงแหวน M ที่สร้างขึ้นใน CPM ของเซลล์ ดังนั้นลักษณะโครงสร้างของฐานของแฟลเจลลัมจึงถูกกำหนดโดยโครงสร้างของ CS
ตามหน้าที่ ตัวฐานเป็นมอเตอร์ไฟฟ้าที่ขับเคลื่อนโดยโปรตอน M-ring ของฐาน (rotating rotor) ล้อมรอบด้วยโปรตีนเมมเบรนที่มีประจุลบ (motor stator) เซลล์แบคทีเรียมีกลไกที่มีประสิทธิภาพในการเปลี่ยนพลังงานไฟฟ้าเคมีเป็นพลังงานกล ดังนั้น แบคทีเรียจึงใช้พลังงานประมาณ 0.1% ของพลังงานทั้งหมดที่ใช้ไปกับการทำงานของแฟลเจลลัม เมื่อแฟลเจลลัมทำงาน แรงกระตุ้นของโปรตอนจะถูกใช้ ซึ่งได้มาจากความแตกต่างของความเข้มข้นของโปรตอนที่ด้านนอกและด้านในของเมมเบรน (มีมากกว่าที่ด้านนอก) และการมีประจุลบมากกว่า ที่ด้านในของเมมเบรน แรงกระตุ้นของโปรตอนบังคับให้โปรตอนเคลื่อนตัวผ่านตัวฐานเข้าไปในเซลล์ ในขณะที่พวกมันยังคงอยู่ในบางส่วนของโรเตอร์ ทำให้เกิดประจุบวก จากนั้นโปรตอนจะเข้าไปในเซลล์ ส่วนที่ชาร์จอยู่ในลักษณะที่แรงดึงดูดเกิดขึ้นระหว่างส่วนที่ชาร์จของโรเตอร์และสเตเตอร์ M-ring เริ่มหมุนด้วยความเร็วประมาณ 300 รอบต่อนาที กลไกการหมุน: การเติมประจุของกลุ่ม COOH ในกรดอะมิโน เพื่อการปฏิวัติที่สมบูรณ์ของวงแหวน โปรตอน 500–1,000 ตัวจะต้องผ่านตัวฐาน การหมุนของวงแหวน M ผ่านแกนที่เชื่อมต่ออย่างแน่นหนากับมัน และขอเกี่ยวจะถูกส่งไปยังเส้นใยแฟลเจลลัม ซึ่งทำหน้าที่เป็นใบพัดหรือใบพัดของเรือ แบคทีเรียจะว่ายตราบใดที่สกรูทำงาน การมีส่วนร่วมของแรงเฉื่อยมีน้อยมาก
นอกจากนี้ แบคทีเรีย แม้แต่แบคทีเรียที่ตายแล้ว ในสภาพแวดล้อมทางน้ำก็เคลื่อนที่อันเป็นผลมาจากการเคลื่อนที่แบบบราวเนียน เซลล์แบคทีเรียต้องอยู่ภายใต้ผลกระทบของโมเลกุลที่อยู่รอบข้างตลอดเวลา ซึ่งอยู่ในการเคลื่อนที่ด้วยความร้อน การโจมตีจากทิศทางต่าง ๆ โยนแบคทีเรียจากทางด้านข้าง
ประเภทของการเคลื่อนที่ของแฟลกเจลลาเป็นแบบหมุน การเคลื่อนไหวมีสองประเภท: เป็นเส้นตรงและไม้ลอย (การเปลี่ยนแปลงแบบสุ่มเป็นระยะในทิศทางของการเคลื่อนไหว) เมื่อแฟลกเจลลาหมุนทวนเข็มนาฬิกา (ประมาณ 1 วินาที) ที่ความถี่ 40-60 รอบต่อนาที (ใกล้เคียงกับความเร็วของมอเตอร์ไฟฟ้าทั่วไป) ด้ายของพวกมันจะถูกทอเป็นมัดเดียว (รูปที่ 9a) การหมุนของแฟลกเจลลาจะถูกส่งไปยังเซลล์ เนื่องจากเซลล์มีขนาดใหญ่กว่าแฟลเจลลัมมาก มันจึงเริ่มเคลื่อนที่เป็นเส้นตรงในทิศทางตรงกันข้าม ด้วยความเร็วที่น้อยกว่าความเร็วของแฟลเจลลัมถึง 3 เท่า
สิ่งนี้ทำให้มั่นใจถึงการเคลื่อนที่ของการแปลของเซลล์ ซึ่งความเร็วของตัวกลางที่เป็นของเหลวสำหรับ ประเภทต่างๆแบคทีเรียอยู่ที่ 20–200 μm/s (ซึ่งสอดคล้องกับความยาวลำตัวประมาณ 300–3000 ต่อนาที) และเคลื่อนไหวช้าลงบนพื้นผิวของตัวกลางที่เป็นของแข็ง
แบคทีเรียสามารถว่ายน้ำอย่างตั้งใจในทิศทางเดียวได้ไม่เกิน 3 วินาที จากนั้นผลกระทบของโมเลกุลโดยรอบจะเปลี่ยนทิศทางแบบสุ่ม ดังนั้นแบคทีเรียจึงได้พัฒนากลไกสำหรับการเปลี่ยนแปลงที่เกิดขึ้นเองในทิศทางของการเคลื่อนไหว - การเปลี่ยนแฟลเจลลาร์มอเตอร์ เมื่อมันเริ่มหมุนตามเข็มนาฬิกา (ประมาณ 0.1 วินาที) แบคทีเรียจะหยุดและพลิกกลับ (ทำการ "กลิ้ง") ไปในทิศทางแบบสุ่ม ในกรณีนี้ แฟลกเจลลาจะกระจายไปในทิศทางต่างๆ (รูปที่ 9b) ในสะเทินน้ำสะเทินบก เมื่อเคลื่อนที่ แฟลกเจลลามัดหนึ่งจะกลับเข้าด้านในออก (เหมือนร่มที่ลมพัดเข้ามา) จากนั้นมอเตอร์จะเปลี่ยนเป็นการหมุนทวนเข็มนาฬิกาอีกครั้ง และแบคทีเรียก็ว่ายเป็นเส้นตรงอีกครั้ง แต่ในทิศทางที่ต่างออกไป
แฟลกเจลลายังสามารถเปลี่ยนทิศทางเพื่อตอบสนองต่อสิ่งเร้าภายนอก หากแบคทีเรียเคลื่อนที่ไปสู่ความเข้มข้นของสารดึงดูดที่เหมาะสม แฟลกเจลลาจะดันเซลล์ผ่านตัวกลาง การเคลื่อนที่เป็นเส้นตรงจะยาวขึ้น และความถี่ในการเคลื่อนตัวลดลง ซึ่งจะทำให้เซลล์เคลื่อนที่ไปในทิศทางที่ต้องการได้ในที่สุด
กรณีของการมีอยู่ของแฟลกเจลลาที่ไม่ใช้งาน (อัมพาต) เป็นที่ทราบกันดีอยู่แล้ว สำหรับการเคลื่อนที่ของแบคทีเรียที่ติดแฟลกเจลนั้น จำเป็นต้องมีความสมบูรณ์ (ไม่เสียหาย) ของ CS การรักษาเซลล์ด้วยไลโซไซม์ ซึ่งนำไปสู่การกำจัดชั้น peptidoglycan ของ CS ทำให้สูญเสียความสามารถในการเคลื่อนที่ของแบคทีเรียแม้ว่าแฟลกเจลลาจะยังคงอยู่
แท็กซี่แบคทีเรีย.บาย สิ่งแวดล้อมยังคงไม่เปลี่ยนแปลง แบคทีเรียว่ายน้ำแบบสุ่ม อย่างไรก็ตาม สภาพแวดล้อมนั้นแทบจะไม่มีความเป็นเนื้อเดียวกันเลย หากสภาพแวดล้อมต่างกัน แบคทีเรียจะแสดงปฏิกิริยาเชิงพฤติกรรมเบื้องต้น: พวกมันจะเคลื่อนที่ไปในทิศทางที่กำหนดโดยปัจจัยภายนอกบางอย่าง การเคลื่อนไหวของแบคทีเรียที่มีจุดประสงค์ทางพันธุกรรมดังกล่าวเรียกว่าแท็กซี่ ขึ้นอยู่กับปัจจัย chemotaxis (กรณีพิเศษคือ aerotaxis), phototaxis, magnetotaxis, thermotaxis และ viscositaxis ขึ้นอยู่กับปัจจัย
เคมีบำบัด- เคลื่อนที่ไปในทิศทางที่แน่นอนเมื่อเทียบกับแหล่งที่มาของสารเคมี สารเคมีแบ่งออกเป็นสองกลุ่ม: เฉื่อยและก่อให้เกิดแท็กซี่ - เคมีบำบัด ในบรรดาสารเคมีที่มีปฏิกิริยาเคมี มีสารที่ดึงดูดแบคทีเรีย - สารดึงดูด (น้ำตาล, กรดอะมิโน, วิตามิน, นิวคลีโอไทด์) และสารที่ขับไล่พวกมัน - สารขับไล่ (กรดอะมิโนบางชนิด, แอลกอฮอล์, ฟีนอล, อิออนอนินทรีย์) สารดึงดูดสำหรับแอโรบิกและสารขับไล่สำหรับโปรคาริโอตที่ไม่ใช้ออกซิเจนคือออกซิเจนระดับโมเลกุล สารดึงดูดมักจะเป็นตัวแทนจากพื้นผิวอาหาร แม้ว่าสารทั้งหมดที่จำเป็นสำหรับร่างกายจะไม่ทำหน้าที่เป็นตัวดึงดูด นอกจากนี้ สารพิษบางชนิดไม่ได้ทำหน้าที่เป็นสารขับไล่และไม่ใช่สารขับไล่ทุกชนิดที่เป็นอันตราย ดังนั้นแบคทีเรียจึงไม่สามารถทำปฏิกิริยากับสารประกอบใด ๆ ได้ แต่เฉพาะกับบางชนิดและต่างกันสำหรับแบคทีเรียที่แตกต่างกัน
ในโครงสร้างพื้นผิวของเซลล์แบคทีเรียมีโมเลกุลโปรตีนพิเศษ - ตัวรับที่เชื่อมต่อกับสารเคมีบางชนิดโดยเฉพาะในขณะที่โมเลกุลของสารเคมีไม่เปลี่ยนแปลง แต่การเปลี่ยนแปลงโครงสร้างเกิดขึ้นในโมเลกุลของตัวรับ ตัวรับจะตั้งอยู่ไม่เท่ากันทั่วทั้งพื้นผิวของเซลล์ และกระจุกตัวอยู่ที่เสาใดขั้วหนึ่ง สถานะของตัวรับสะท้อนถึงความเข้มข้นนอกเซลล์ของเอฟเฟกต์ที่สอดคล้องกัน
Chemotaxis มีค่าปรับตัว ตัวอย่างเช่น รูปแบบของอหิวาตกโรค วิบริโอที่มีเคมีบำบัดบกพร่องจะมีความรุนแรงน้อยกว่า
แอโรแทกซิส- แบคทีเรียที่ต้องการอ๊อกซิเจนระดับโมเลกุลจะสะสมอยู่รอบๆ ฟองอากาศที่ตกอยู่ใต้ผ้าคลุม
โฟโต้แท็กซี่- การเคลื่อนที่เข้าหาหรือออกจากแสง ลักษณะของแบคทีเรีย phototrophic ที่ใช้แสงเป็นแหล่งพลังงาน
แมกนีโตแทกซิส- ความสามารถของแบคทีเรียในน้ำที่มีผลึกของแร่ธาตุที่มีธาตุเหล็กว่ายตามแนวสนามแม่เหล็กของโลก
เทอร์โมแทกซิส- การเคลื่อนที่ไปในทิศทางของการเปลี่ยนแปลงของอุณหภูมิซึ่งมี สำคัญมากสำหรับแบคทีเรียก่อโรคบางชนิด
Viscositaxis- ความสามารถในการตอบสนองต่อการเปลี่ยนแปลงความหนืดของสารละลาย แบคทีเรียมักจะมีแนวโน้มเป็นสื่อที่มีความหนืดสูงกว่า ซึ่งมีความสำคัญอย่างยิ่งสำหรับสายพันธุ์ที่ทำให้เกิดโรค
แบคทีเรียเลื่อน.ความสามารถในการเลื่อนที่ความเร็วต่ำ (2–11 µm/s) บนซับสเตรตที่เป็นของแข็งหรือหนืดนั้นพบได้ในโปรคาริโอตบางชนิด เช่น มัยโคพลาสมา
มีสมมติฐานหลายประการที่อธิบายการเคลื่อนที่แบบเลื่อน ตาม สมมติฐานการขับเคลื่อนด้วยไอพ่นเกิดจากการหลั่งเมือกผ่านรูพรุนของเมือกจำนวนมากใน SC ซึ่งเป็นผลมาจากการที่เซลล์ถูกขับออกจากสารตั้งต้นไปในทิศทางตรงกันข้ามกับทิศทางของการหลั่งเมือก ตาม สมมติฐานคลื่นเดินทางการเคลื่อนที่แบบร่อนในรูปแบบที่ปราศจากแฟลเจลเลตที่เคลื่อนที่ได้สัมพันธ์กับการมีอยู่ระหว่างชั้น peptidoglycan และเยื่อหุ้มชั้นนอกของ CL ของชั้นโปรตีนบางๆ ของไฟบริลสั่งการ คล้ายกับเส้นใยของแฟลเจลลา การเคลื่อนที่แบบหมุนของเส้นใยที่ "เปิด" โดยโครงสร้างเหล่านี้ นำไปสู่การปรากฏตัวของ "คลื่นเคลื่อนที่" (การเคลื่อนตัวนูนด้วยกล้องจุลทรรศน์ของ CL) บนผิวเซลล์ ซึ่งเป็นผลมาจากการที่เซลล์ถูกขับออกจากสารตั้งต้น ในที่สุด โครงสร้างที่คล้ายกับฐานของรูปแบบแฟลเจลได้อธิบายไว้ในแบคทีเรียร่อนบางชนิด
หน้าที่ของแฟลกเจลลา:
1. ให้การยึดเกาะ - ชั้นต้นกระบวนการติดเชื้อ
2. ให้การเคลื่อนไหวของแบคทีเรีย
3. กำหนดความจำเพาะของแอนติเจน นี่คือ H-antigen
การตรวจหาแฟลกเจลลา:
1. กล้องจุลทรรศน์เฟสคอนทราสต์ของการเตรียมพื้นเมือง (“ บดขยี้” และ“ ห้อย”) ด้วยกล้องจุลทรรศน์ การเคลื่อนที่จะถูกกำหนดในเซลล์ของวัฒนธรรมประจำวัน เพื่อแยกความแตกต่างของการเคลื่อนไหวออกจากการเคลื่อนที่แบบบราวเนียนแบบพาสซีฟ หยดสารละลายฟีนอล 5% ในน้ำจะถูกเติมลงในวัฒนธรรมที่ศึกษาหนึ่งหยด ในกรณีนี้ การเคลื่อนไหวแบบแอคทีฟจะหยุดลง
2. กล้องจุลทรรศน์สนามมืดของการเตรียมพื้นเมือง
3. กล้องจุลทรรศน์แสงของการเตรียมการย้อมด้วยสีย้อมหรือโลหะ เนื่องจากแฟลกเจลลาเกิดความเสียหายได้ง่ายมากระหว่างการเตรียมการ วิธีการเหล่านี้จึงไม่ค่อยได้ใช้ในชีวิตประจำวัน
แฟลกเจลลาถูกย้อมโดยใช้เซลล์ที่ปลูกบนแนวเอียงของวุ้น เซลล์ที่อยู่ใกล้น้ำกลั่นตัวจะถูกเลือกด้วยวงแบคทีเรียและย้ายอย่างระมัดระวังไปยังน้ำกลั่นที่ปราศจากเชื้อซึ่งมีอุณหภูมิเท่ากันกับอุณหภูมิของการฟักตัวของแบคทีเรียบนวุ้นลาดเอียง และแบคทีเรียจะไม่ถูกสะบัดออกจากวงจร แต่จะจุ่มลงในน้ำอย่างระมัดระวัง หลอดที่มีแบคทีเรียถูกทิ้งไว้ที่อุณหภูมิห้องเป็นเวลา 30 นาที ใช้แก้วบริสุทธิ์ทางเคมี (ล้างด้วยส่วนผสมโครเมียม) โดยใช้สารแขวนลอย 2-3 หยด ระบบกันสะเทือนจะกระจายไปทั่วพื้นผิวกระจกโดยค่อยๆ เอียงมัน ตากยาให้แห้ง
แฟลกเจลลานั้นบางมาก จึงสามารถตรวจพบได้ด้วยกระบวนการพิเศษเท่านั้น ในขั้นต้นด้วยความช่วยเหลือของการแกะสลักการบวมและการเพิ่มขนาดของพวกเขาทำได้จากนั้นการเตรียมการจะเป็นสีเนื่องจากมองเห็นได้ภายใต้กล้องจุลทรรศน์แบบใช้แสง
ใช้บ่อยขึ้น วิธีการทำเงินตาม Morozov (รูปที่ 10):
- การเตรียมได้รับการแก้ไขด้วยสารละลายกรดอะซิติกน้ำแข็งเป็นเวลา 1 นาทีล้างด้วยน้ำ
- ใช้สารละลายแทนนิน (ฟอกทำให้แฟลเจลลามีความหนาแน่นมากขึ้น) เป็นเวลา 1 นาทีล้างด้วยน้ำ
- การเตรียมการรักษาโดยการให้ความร้อนด้วยสารละลายซิลเวอร์ไนเตรต 1-2 นาทีล้างด้วยน้ำแห้งและกล้องจุลทรรศน์
ด้วยกล้องจุลทรรศน์จะมองเห็นเซลล์สีน้ำตาลเข้มและแฟลกเจลลาที่เบากว่า

ข้าว. สิบ.การระบุแฟลกเจลลาโดยการลงเงิน
ข้าว. สิบเอ็ดบัตรประจำตัวของแฟลกเจลลา
โดยกล้องจุลทรรศน์อิเล็กตรอน
มะเดื่อ 4. กล้องจุลทรรศน์อิเล็กตรอนของการเตรียมการที่สะสมด้วยโลหะหนัก (รูปที่ 11)
5. ทางอ้อม - ตามลักษณะของการเจริญเติบโตของแบคทีเรียเมื่อหว่านในวุ้นกึ่งของเหลว 0.3% หลังจากการฟักตัวของพืชในเทอร์โมสตัทเป็นเวลา 1–2 วันจะสังเกตเห็นลักษณะของการเติบโตของแบคทีเรีย:
- ในแบคทีเรียที่ไม่เคลื่อนที่ (เช่น S.saprophyticus) มีการเจริญเติบโตตามการฉีด - "เล็บ" และสื่อมีความโปร่งใส
ในแบคทีเรียที่เคลื่อนที่ได้ (เช่น อี. โคไล) มีการเติบโตที่ด้านข้างของการฉีด ตลอดทั้งคอลัมน์วุ้น - "ก้างปลา" และกระจายความขุ่นของตัวกลาง
แบคทีเรียทั้งหมดแบ่งเป็นแบบเคลื่อนที่และไม่เคลื่อนที่ อวัยวะของการเคลื่อนไหวในแบคทีเรียคือแฟลกเจลลา ประกอบด้วยแฟลเจลลินซึ่งเป็นโปรตีนที่มีโครงสร้างเกี่ยวข้องกับโปรตีนหดตัวประเภทไมโอซิน
ฐานแฟลเจลลัมเป็นร่างกายฐานซึ่งประกอบด้วยระบบของดิสก์ (blepharoplast: 1 ดิสก์ - ด้านนอกของผนังเซลล์, 2 ดิสก์ - ด้านในของผนังเซลล์, 3 ดิสก์ - เยื่อหุ้มเซลล์ไซโตพลาสซึม), "ฝัง" ในไซโตพลาสซึม เยื่อหุ้มเซลล์และผนังเซลล์ ความยาวของแฟลเจลลัมนั้นมากกว่าความยาวของลำตัวของจุลินทรีย์เอง
ตามจำนวนแฟลกเจลลาและที่ตั้งของจุลินทรีย์เคลื่อนที่แบ่งออกเป็น:
1. Monotrichous มีแฟลเจลลัมหนึ่งตัวที่ส่วนท้ายของร่างกาย (เคลื่อนที่ได้มากที่สุด) ตัวอย่างเช่น Vibrio cholerae
2. Lophotrichous มีแฟลกเจลลามัดหนึ่งมัดที่ขั้วใดขั้วหนึ่งของเซลล์ ตัวอย่างเช่น Burkholderia (Pseudomonas) pseudomalei เป็นสาเหตุของโรคเมลิออยด์
3. Amphitrichous มีแฟลเจลลัมอยู่ที่ขั้วทั้งสองเซลล์ ตัวอย่างเช่น Spirillum volutans
4. Peritrichous มีแฟลกเจลลารอบปริมณฑลของเซลล์ ตัวอย่างเช่น Escherichia coli, Salmonella typhi
การตรวจหาแฟลกเจลลาแฟลกเจลลานั้นบางมาก จึงสามารถตรวจพบได้ด้วยกระบวนการพิเศษเท่านั้น โดยเฉพาะอย่างยิ่งในตอนแรกด้วยความช่วยเหลือของ mordant อาการบวมและการเพิ่มขนาดของพวกเขาจะทำได้และจากนั้นการเตรียมการจะเป็นสีเนื่องจากสามารถมองเห็นได้ภายใต้กล้องจุลทรรศน์แบบใช้แสง สามารถตรวจพบแฟลกเจลลาได้โดยการย้อมสีตาม Morozov, Leffler และด้วยกล้องจุลทรรศน์อิเล็กตรอน แฟลกเจลลายังสามารถตรวจพบได้จากการเคลื่อนไหวของแบคทีเรีย
สังเกตการเคลื่อนไหวของจุลินทรีย์ในการเตรียมหยด "บด" และ "ห้อย" จากวัฒนธรรมที่มีชีวิต การเตรียมเหล่านี้ใช้กล้องจุลทรรศน์ด้วยเลนส์แบบแห้งหรือแบบจุ่มในที่มืดหรือแบบเฟสคอนทราสต์ นอกจากนี้ การเคลื่อนที่สามารถกำหนดได้โดยธรรมชาติของการเติบโตของแบคทีเรียในวุ้นกึ่งของเหลว
ดื่มจากแบคทีเรีย
Pili (pili) คำพ้องความหมาย: villi, fimbria เป็นเส้นใยกลวงบาง ๆ ที่มีลักษณะโปรตีนซึ่งครอบคลุมพื้นผิวของเซลล์แบคทีเรีย ต่างจากแฟลเจลลาตรงที่พวกมันไม่ทำหน้าที่เกี่ยวกับมอเตอร์
Pili ขยายออกจากผิวเซลล์และประกอบด้วยโปรตีน pilina.
ตามวัตถุประสงค์การใช้งานจะแบ่งออกเป็น 2 ประเภท
1) Pili ชนิดแรกมีอยู่ในแบคทีเรียส่วนใหญ่จึงเรียกว่า "villi ชนิดทั่วไป" (common pili) ทำให้เกิดการยึดติดหรือการยึดเกาะของแบคทีเรียกับเซลล์บางชนิดของสิ่งมีชีวิตที่เป็นโฮสต์ การยึดเกาะเป็นขั้นตอนเริ่มต้นของกระบวนการติดเชื้อใดๆ
2) Pili ประเภทที่สอง (คำพ้องความหมาย: conjugative หรือ sexual - sex pili) มีให้เฉพาะในแบคทีเรียผู้บริจาคที่มีพลาสมิดพิเศษ จำนวนของพวกเขามีขนาดเล็ก - 1-4 ต่อเซลล์
เลื่อยพื้นทำหน้าที่ดังต่อไปนี้:
1. มีส่วนร่วมในการถ่ายโอนสารพันธุกรรมจากเซลล์หนึ่งไปยังอีกเซลล์หนึ่งในระหว่างการผันของแบคทีเรีย
2. ดูดซับไวรัสแบคทีเรียจำเพาะ - bacteriophages
สปอร์ของแบคทีเรีย สภาพการก่อตัว ตำแหน่ง กลไกและระยะของการย้อมสีเอาเจสกี้
การโต้เถียง- รูปแบบแปลก ๆ ของแบคทีเรียที่พักผ่อนโดยมีโครงสร้างผนังเซลล์แบบแกรมบวก
การสร้างสปอร์- นี่เป็นวิธีการรักษาสายพันธุ์ (genophore) ในสภาพแวดล้อมภายนอกภายใต้สภาวะที่ไม่เอื้ออำนวยและไม่ใช่วิธีการสืบพันธุ์
สปอร์ก่อตัวขึ้นภายใต้สภาวะที่ไม่เอื้ออำนวยต่อการดำรงอยู่ของแบคทีเรีย (การทำให้แห้ง การขาดสารอาหาร ฯลฯ) ภายในเซลล์แบคทีเรียจะเกิดสปอร์หนึ่งตัว (เอนโดสปอร์)
ขั้นตอนของการสร้างสปอร์
1. การเตรียมการ ในไซโตพลาสซึมของแบคทีเรีย บริเวณที่อัดแน่นจะก่อตัวขึ้นซึ่งไม่มีน้ำอิสระ เรียกว่า "โซนสปอโรจีนัส" ซึ่งมีนิวเคลียสอยู่
2. ระยะของพรีสปอร์ (prospores) รอบเขต sporogenous จะเกิดเปลือกของเยื่อหุ้มเซลล์ไซโตพลาสซึมสองชั้น
3. การก่อตัวของเยื่อหุ้มสมองที่ประกอบด้วย peptidoglycan และเยื่อหุ้มชั้นนอกที่มีเกลือแคลเซียมและไขมันสูง
4. ขั้นตอนของการเจริญเติบโต ซองสปอร์จะก่อตัวขึ้นที่ด้านนอกของเยื่อหุ้มชั้นนอก หลังจากนั้นส่วนที่เป็นพืชของเซลล์จะถูกสลาย ปล่อยสปอร์ออกมา
การพัฒนาทางจุลชีววิทยาทำให้เกิดการค้นพบมากมายในทศวรรษที่ผ่านมา และหนึ่งในนั้นคือลักษณะเฉพาะของการเคลื่อนไหวของแบคทีเรียที่ติดธง การออกแบบเครื่องยนต์ของสิ่งมีชีวิตโบราณเหล่านี้กลายเป็นเรื่องที่ซับซ้อนมากและตามหลักการทำงานของพวกมันนั้นแตกต่างจากแฟลกเจลลาของญาติของยูคาริโอตที่ใกล้เคียงที่สุดของโปรโตซัว เครื่องยนต์ของแบคทีเรียแฟลเจลเลตเป็นประเด็นถกเถียงที่ร้อนแรงที่สุดระหว่างผู้ทรงสร้างและนักวิวัฒนาการ เกี่ยวกับแบคทีเรีย มอเตอร์แฟลกเจลลาร์ และอื่นๆ อีกมากมาย - บทความนี้
ชีววิทยาทั่วไป
ในการเริ่มต้น ให้เราระลึกว่าพวกมันเป็นสิ่งมีชีวิตประเภทใดและพวกมันอาศัยอยู่ที่ใดในระบบของโลกอินทรีย์บนโลกของเรา โดเมนแบคทีเรียรวมสิ่งมีชีวิตโปรคาริโอตที่มีเซลล์เดียวจำนวนมาก (โดยไม่มีนิวเคลียสที่ก่อตัวขึ้น) เข้าด้วยกัน
เซลล์ที่มีชีวิตเหล่านี้ปรากฏขึ้นในที่เกิดเหตุเมื่อเกือบ 4 พันล้านปีก่อน และเป็นผู้ตั้งถิ่นฐานกลุ่มแรกในโลก พวกมันสามารถมีรูปร่างที่แตกต่างกันมาก (cocci, rods, vibrios, spirochetes) แต่ส่วนใหญ่จะถูกแฟลกเจลลา
แบคทีเรียอาศัยอยู่ที่ไหน? ทุกที่. มากกว่า 5 × 10 30 อาศัยอยู่บนโลก มีประมาณ 40 ล้านตัวในดิน 1 กรัม มากถึง 39 ล้านล้านอาศัยอยู่ในร่างกายของเรา พวกมันสามารถพบได้ที่ก้นร่องลึกบาดาลมาเรียนา ใน "กลุ่มควันดำ" ที่ร้อนระอุที่ก้นมหาสมุทร ในน้ำแข็งของทวีปแอนตาร์กติกา และขณะนี้คุณมีแบคทีเรียมากถึง 10 ล้านตัวในมือของคุณ
คุณค่าที่ปฏิเสธไม่ได้
แม้จะมีขนาดจุลทรรศน์ (0.5-5 ไมครอน) ชีวมวลรวมของพวกมันบนโลกนั้นยิ่งใหญ่กว่าชีวมวลของสัตว์และพืชรวมกัน บทบาทของพวกเขาในการไหลเวียนของสารไม่สามารถถูกแทนที่ได้และคุณสมบัติของผู้บริโภค (ผู้ทำลายสารอินทรีย์) ไม่อนุญาตให้โลกถูกปกคลุมด้วยซากศพ
อย่าลืมเกี่ยวกับเชื้อโรค: สาเหตุของกาฬโรค, ไข้ทรพิษ, ซิฟิลิส, วัณโรคและโรคติดเชื้ออื่น ๆ อีกมากมายก็เป็นแบคทีเรียเช่นกัน
แบคทีเรียได้พบการประยุกต์ใช้ในกิจกรรมทางเศรษฐกิจของมนุษย์ เริ่มจากอุตสาหกรรมอาหาร ( ผลิตภัณฑ์นม, ชีส, ผักดอง, เครื่องดื่มแอลกอฮอล์) เศรษฐกิจ "สีเขียว" (เชื้อเพลิงชีวภาพและก๊าซชีวภาพ) ต่อวิธีวิศวกรรมเซลล์และการผลิตยา (วัคซีน เซรั่ม ฮอร์โมน วิตามิน)

สัณฐานวิทยาทั่วไป
ดังที่ได้กล่าวไปแล้วตัวแทนของสิ่งมีชีวิตที่มีเซลล์เดียวเหล่านี้ไม่มีนิวเคลียสวัสดุทางพันธุกรรมของพวกมัน (โมเลกุล DNA ในรูปของวงแหวน) ตั้งอยู่ในพื้นที่บางส่วนของไซโตพลาสซึม เซลล์ของพวกมันมีพลาสมาเมมเบรนและแคปซูลหนาแน่นที่เกิดจาก peptidoglycan murein ในออร์แกเนลล์ของเซลล์ แบคทีเรียมีไมโตคอนเดรีย อาจมีคลอโรพลาสต์และโครงสร้างอื่นๆ ที่มีหน้าที่ต่างกัน
แบคทีเรียส่วนใหญ่เป็นแฟลกเจลลา แคปซูลที่แน่นบนพื้นผิวของเซลล์ป้องกันไม่ให้พวกมันเคลื่อนที่ไปรอบๆ โดยการเปลี่ยนตัวเซลล์เอง เช่นเดียวกับอะมีบา แฟลกเจลลาของพวกมันคือการก่อตัวของโปรตีนหนาแน่นที่มีความยาวต่างกันและมีเส้นผ่านศูนย์กลางประมาณ 20 นาโนเมตร แบคทีเรียบางชนิดมีแฟลเจลลัมเดี่ยว (monotrichous) ในขณะที่บางชนิดมีแฟลเจลลัมสองตัว (amphitrichous) บางครั้งแฟลเจลลาจะถูกจัดเรียงเป็นมัด (lophorichous) หรือครอบคลุมพื้นผิวทั้งหมดของเซลล์ (peritrichous)
พวกมันจำนวนมากอาศัยอยู่เป็นเซลล์เดียว แต่มีบางรูปแบบเป็นกระจุก (คู่, โซ่, เส้นใย, เส้นใย)

คุณสมบัติการเคลื่อนไหว
แบคทีเรียที่ติดธงสามารถเคลื่อนที่ได้หลายวิธี บางคนก้าวไปข้างหน้าเท่านั้น และเปลี่ยนทิศทางด้วยการล้มลง บางตัวสามารถกระตุกได้ บางตัวสามารถขยับได้โดยการเลื่อน
แฟลกเจลลาของแบคทีเรียทำหน้าที่ไม่เพียง แต่ "พาย" ในระดับเซลล์เท่านั้น แต่ยังสามารถเป็นเครื่องมือ "ขึ้นเครื่อง" ได้อีกด้วย
จนกระทั่งเมื่อไม่นานมานี้ เชื่อกันว่าแฟลเจลลัมของแบคทีเรียจะกระดิกหางเหมือนงู การศึกษาเมื่อเร็ว ๆ นี้แสดงให้เห็นว่าแฟลเจลลัมของแบคทีเรียนั้นซับซ้อนกว่ามาก มันทำงานเหมือนกังหัน ติดกับไดรฟ์จะหมุนไปในทิศทางเดียว ตัวกระตุ้นหรือมอเตอร์แฟลเจลลาร์ของแบคทีเรียเป็นโครงสร้างโมเลกุลที่ซับซ้อนซึ่งทำงานเหมือนกล้ามเนื้อ ด้วยความแตกต่างที่กล้ามเนื้อต้องคลายตัวหลังจากการหดตัวและมอเตอร์ของแบคทีเรียก็ทำงานอย่างต่อเนื่อง

กลไกนาโนของแฟลเจลลัม
โดยไม่ต้องเจาะลึกถึงชีวเคมีของการเคลื่อนไหว เราสังเกตว่ามีโปรตีนมากถึง 240 ตัวที่เกี่ยวข้องกับการสร้างแฟลเจลลัมไดรฟ์ ซึ่งแบ่งออกเป็นส่วนประกอบระดับโมเลกุล 50 ตัวที่มีหน้าที่เฉพาะในระบบ
ในระบบขับเคลื่อนของแบคทีเรียนี้มีโรเตอร์ที่เคลื่อนที่และสเตเตอร์ที่ให้การเคลื่อนไหวนี้ มีเพลาขับ บุชชิ่ง คลัตช์ เบรค และคันเร่ง
เครื่องยนต์จิ๋วนี้ช่วยให้แบคทีเรียเดินทางได้ 35 เท่าของขนาดตัวของมันเองในเวลาเพียง 1 วินาที ในเวลาเดียวกันการทำงานของแฟลเจลลัมเองซึ่งทำ 60,000 รอบต่อนาทีร่างกายใช้พลังงานเพียง 0.1% ของพลังงานทั้งหมดที่เซลล์ใช้ไป
ยังเป็นที่น่าประหลาดใจอีกด้วยที่แบคทีเรียสามารถแทนที่และซ่อมแซมชิ้นส่วนอะไหล่ทั้งหมดของกลไกมอเตอร์ของมันได้ "ในระหว่างเดินทาง" แค่คิดว่าคุณอยู่บนเครื่องบิน และช่างเปลี่ยนใบพัดของมอเตอร์ที่กำลังทำงานอยู่
แบคทีเรียแฟลกเจลเลตกับดาร์วิน
เครื่องยนต์ที่สามารถทำงานได้ด้วยความเร็วสูงถึง 60,000 รอบต่อนาที สตาร์ทด้วยตนเองและใช้คาร์โบไฮเดรต (น้ำตาล) เป็นเชื้อเพลิงเท่านั้น มีอุปกรณ์ที่คล้ายกับมอเตอร์ไฟฟ้า อุปกรณ์ดังกล่าวสามารถเกิดขึ้นได้ในกระบวนการวิวัฒนาการหรือไม่?
นี่คือคำถามที่ Michael Behe, Ph.D. ในสาขาชีววิทยา ถามตัวเองในปี 1988 เขาแนะนำแนวคิดเกี่ยวกับระบบที่ลดไม่ได้ในระบบชีววิทยาซึ่งเป็นระบบที่ทุกส่วนของมันจำเป็นพร้อม ๆ กันเพื่อให้แน่ใจว่ามีการทำงานและการกำจัดอย่างน้อยหนึ่งส่วนทำให้เกิดการหยุดชะงักของการทำงานอย่างสมบูรณ์
จากจุดยืนของวิวัฒนาการของดาร์วิน การเปลี่ยนแปลงโครงสร้างทั้งหมดในร่างกายจะเกิดขึ้นทีละน้อย และมีเพียงการเปลี่ยนแปลงที่ประสบความสำเร็จเท่านั้นที่ถูกคัดเลือกโดยการคัดเลือกโดยธรรมชาติ
บทสรุปของ M. Behe ระบุไว้ในหนังสือ "Darwin's Black Box" (1996): เครื่องยนต์ของแฟลเจลลาร์แบคทีเรียเป็นระบบที่แบ่งแยกไม่ได้มากกว่า 40 ส่วน และการไม่มีอย่างน้อยหนึ่งส่วนจะทำให้เกิดความผิดปกติอย่างสมบูรณ์ของ ระบบซึ่งหมายความว่าระบบนี้ไม่สามารถเกิดขึ้นได้โดยการคัดเลือกโดยธรรมชาติ

บาล์มสำหรับนักสร้างสรรค์
ทฤษฎีการสร้างสรรค์ที่นำเสนอโดยนักวิทยาศาสตร์และศาสตราจารย์ด้านชีววิทยาคณบดีคณะวิทยาศาสตร์ชีวภาพที่ Lehigh University of Bethlehem (USA) M. Behe ดึงดูดความสนใจของรัฐมนตรีคริสตจักรและผู้สนับสนุนทฤษฎีของพระเจ้าในทันที ต้นกำเนิดของชีวิต
ในปี 2548 ในสหรัฐอเมริกายังมีคดีที่ Behe เป็นพยานจากผู้สนับสนุนทฤษฎี "การออกแบบที่สมเหตุสมผล" ซึ่งถือเป็นการแนะนำการศึกษาเรื่องการเนรเทศในโรงเรียนโดเวอร์ในหลักสูตร " เกี่ยวกับแพนด้าและผู้คน” กระบวนการนี้สูญหายไปการสอนเรื่องดังกล่าวได้รับการยอมรับว่าขัดต่อรัฐธรรมนูญฉบับปัจจุบัน
แต่การโต้เถียงระหว่างผู้สร้างโลกและนักวิวัฒนาการยังคงดำเนินต่อไปในทุกวันนี้
แฟลเจลลัมเป็นโครงสร้างพื้นผิวของเซลล์แบคทีเรีย ซึ่งทำหน้าที่ในการเคลื่อนที่ในตัวกลางที่เป็นของเหลว
ขึ้นอยู่กับตำแหน่งของแฟลกเจลลา แบคทีเรียแบ่งออกเป็น (รูปที่ 1):
Peritrichial
ผสม
เสา
Subpolar
เสาแฟลเจลลา- แฟลกเจลลาตั้งแต่หนึ่งตัวขึ้นไปตั้งอยู่บนขั้วเดียว (โมโนโพลาร์) หรือทั้งสองขั้ว (ไบโพลาร์) ของเซลล์ และฐานขนานกับแกนยาวของเซลล์
แฟลกเจลลาใต้ขั้ว(subpolar) - แฟลกเจลลาตั้งแต่หนึ่งตัวขึ้นไปอยู่ที่จุดเปลี่ยนของพื้นผิวด้านข้างไปยังขั้วของเซลล์ที่ปลายหนึ่งหรือสองด้าน ที่ฐานเป็นมุมฉากกับแกนยาวของเซลล์
แฟลกเจลลาด้านข้าง(ด้านข้าง) - แฟลกเจลลาหนึ่งตัวหรือมากกว่าในรูปแบบของมัดตั้งอยู่ที่จุดกึ่งกลางของครึ่งหนึ่งของเซลล์
แฟลกเจลลาในช่องท้อง- ตั้งอยู่เหนือพื้นผิวทั้งหมดของเซลล์ทีละตัวหรือเป็นกลุ่ม เสามักจะถูกกีดกันออกไป
แฟลกเจลลาผสม- แฟลกเจลลาตั้งแต่สองตัวขึ้นไปอยู่ใน จุดต่างๆเซลล์.
ขึ้นอยู่กับจำนวนของแฟลกเจลลามี:
Monotrichous - หนึ่งแฟลเจลลัม
Polytrichs - พวงแฟลกเจลลา
ยังโดดเด่น:
lophorichous- การจัดเรียง polytrichial แบบผูกขาดของแฟลกเจลลา
amphitriches- การจัดเรียงตัวของแฟลเจลลาสองขั้ว polytrichial
โครงสร้างของแฟลเจลลัมแบคทีเรียและตัวฐาน เฆี่ยน.
แฟลเจลลัมนั้นถูกจัดเรียงอย่างเรียบง่าย: ฟิลาเมนต์ที่ติดอยู่กับตัวฐาน บางครั้งส่วนโค้งของท่อที่เรียกว่าขอเกี่ยวสามารถสอดเข้าไประหว่างตัวฐานกับไส้หลอดได้ มันหนากว่าไส้หลอดและมีส่วนร่วมในการยึดติดที่ยืดหยุ่นของไส้หลอดกับตัวฐาน
ในแง่ขององค์ประกอบทางเคมี แฟลเจลลัมประกอบด้วยโปรตีนแฟลเจลลิน 98% (แฟลเจลลัม - แฟลเจลลัม) ประกอบด้วยกรดอะมิโน 16 ชนิด กลูตามีนและกรดแอสปาร์ติกมีอิทธิพลเหนือ ทริปโตเฟน ซิสเทอีนและซิสทีนขาดกรดอะมิโนอะโรมาติกจำนวนเล็กน้อย แฟลกเจลลินกำหนดความจำเพาะของแอนติเจนเรียกว่า H-antigen แบคทีเรียแฟลกเจลลาไม่มีกิจกรรม ATPase
ความหนาของแฟลเจลลัมคือ 10–12 นาโนเมตร ความยาว 3–15 ไมโครเมตร
เป็นเกลียวแข็งบิดทวนเข็มนาฬิกา การหมุนของแฟลเจลลัมยังดำเนินการทวนเข็มนาฬิกาด้วยความถี่ 40 รอบต่อนาทีถึง 60 รอบต่อนาทีซึ่งทำให้เซลล์หมุนไปในทิศทางตรงกันข้าม แต่เนื่องจาก เซลล์หนักกว่าแฟลเจลลัมมาก จากนั้นการหมุนจะช้าลงจาก 12 ถึง 14 รอบต่อนาที
แฟลเจลลัมเติบโตจากส่วนปลาย โดยที่หน่วยย่อยเข้ามาทางช่องภายใน ในบางสปีชีส์ แฟลเจลลัมยังถูกหุ้มด้านนอกด้วยปลอกหุ้ม ซึ่งเป็นส่วนต่อของผนังเซลล์และอาจมีโครงสร้างเหมือนกัน
ร่างกายพื้นฐาน
ฐานร่างกายประกอบด้วย 4 ส่วน:
ผสมพันธุ์ด้วยด้ายหรือขอเกี่ยว
ดิสก์สองอันพันอยู่บนแท่ง (เอ็มและเอส)
กลุ่มโปรตีนเชิงซ้อน (สเตเตอร์)
โปรตีนแคป
แบคทีเรียที่มีเยื่อหุ้มชั้นในและชั้นนอกมีดิสก์เพิ่มเติม 2 แผ่น (P และ L) และโครงสร้างโปรตีนที่เยื่อหุ้มชั้นนอกใกล้กับตัวฐาน จึงไม่มีบทบาทสำคัญในการเคลื่อนไหว
ลักษณะเฉพาะของโครงสร้างของร่างกายฐานถูกกำหนดโดยโครงสร้างของผนังเซลล์: ความสมบูรณ์ของมันเป็นสิ่งจำเป็นสำหรับการเคลื่อนไหวของแฟลกเจลลา การรักษาเซลล์ด้วยไลโซไซม์นำไปสู่การกำจัดชั้น peptidoglycan ออกจากผนังเซลล์ ซึ่งนำไปสู่การสูญเสียการเคลื่อนไหว แม้ว่าโครงสร้างของแฟลเจลลัมจะไม่ถูกรบกวน
แบคทีเรียแฟลกเจลลากำหนดการเคลื่อนที่ของเซลล์แบคทีเรีย แฟลกเจลลาเป็นเส้นใยบาง ๆ ที่เกิดจากเยื่อหุ้มไซโตพลาสซึมและยาวกว่าตัวเซลล์เอง แฟลกเจลลามีความหนา 12–20 นาโนเมตร และยาว 3–15 ไมโครเมตร ประกอบด้วย 3 ส่วน ได้แก่ เกลียวเกลียว ตะขอ และตัวฐานที่มีแกนพร้อมแผ่นพิเศษ (แผ่นดิสก์ 1 คู่สำหรับแกรมบวกและ 2 คู่สำหรับแบคทีเรียแกรมลบ) แผ่นแฟลกเจลลาติดกับเยื่อหุ้มไซโตพลาสซึมและผนังเซลล์ สิ่งนี้สร้างเอฟเฟกต์ของมอเตอร์ไฟฟ้าด้วยแกนมอเตอร์ที่หมุนแฟลเจลลัม แฟลกเจลลาประกอบด้วยโปรตีน - แฟลเจลลิน (จากแฟลเจลลัม - แฟลเจลลัม); เป็นแอนติเจน H หน่วยย่อยแฟลกเจลลินถูกขด จำนวนแฟลกเจลลาในแบคทีเรียของสปีชีส์ต่างกันแตกต่างกันไปตั้งแต่หนึ่งตัว (โมโนทริช) ใน Vibrio cholerae ถึงสิบหรือหลายร้อยแฟลกเจลลาที่ขยายไปตามขอบของแบคทีเรีย (peritrich) ใน Escherichia coli, Proteus เป็นต้น Lofotrichs มีกลุ่มของ flagella อยู่ที่หนึ่ง ปลายเซลล์ Amphitrichous มีแฟลเจลลัมหนึ่งอันหรือแฟลเจลลามัดหนึ่งมัดที่ปลายอีกด้านของเซลล์
Pili (fimbriae, villi) - การก่อตัวเป็นเส้นใย ทินเนอร์และสั้นกว่า (3-10 nm x 0.3-10 ไมครอน) กว่าแฟลเจลลา Pili ขยายออกจากผิวเซลล์และประกอบด้วยโปรตีน pilin ซึ่งมีฤทธิ์แอนติเจน มี pili ที่มีหน้าที่ในการยึดเกาะ กล่าวคือ สำหรับติดแบคทีเรียกับเซลล์ที่ได้รับผลกระทบ เช่นเดียวกับ pili ที่รับผิดชอบด้านโภชนาการ เมตาบอลิซึมของเกลือน้ำ และทางเพศ (F-pili) หรือพิลีผันคำกริยา เครื่องดื่มมีมากมาย - กรงละหลายร้อยตัว อย่างไรก็ตาม pili เพศมักจะอยู่ที่ 1-3 ต่อเซลล์: พวกมันถูกสร้างขึ้นโดยเซลล์ผู้บริจาคที่เรียกว่า "ผู้ชาย" ที่มีพลาสมิดที่ถ่ายทอดได้ (F-, R-, Col-plasmids) ลักษณะเด่นของ pili เพศคือการมีปฏิสัมพันธ์กับแบคทีเรียทรงกลมพิเศษ "เพศชาย" ซึ่งถูกดูดซับอย่างเข้มข้นบน pili เพศ
สปอร์เป็นรูปแบบเฉพาะของแบคทีเรีย Firmicute ที่อยู่เฉยๆ เช่น แบคทีเรียที่มีโครงสร้างผนังเซลล์แกรมบวก สปอร์เกิดขึ้นภายใต้สภาวะที่ไม่เอื้ออำนวยต่อการดำรงอยู่ของแบคทีเรีย (การทำให้แห้ง การขาดสารอาหาร ฯลฯ สปอร์หนึ่งตัว (เอนโดสปอร์) ก่อตัวขึ้นภายในเซลล์ของแบคทีเรีย การก่อตัวของสปอร์ช่วยรักษาสายพันธุ์และไม่ใช่วิธีการสืบพันธุ์ เช่นในเชื้อรา แบคทีเรียที่สร้างสปอร์ในสกุล Bacillus มีสปอร์ไม่เกินเส้นผ่านศูนย์กลางของเซลล์ แบคทีเรียที่มีขนาดสปอร์เกินเส้นผ่านศูนย์กลางของเซลล์เรียกว่า clostridia เช่น แบคทีเรียในสกุล Clostridium (lat. Clostridium - แกนหมุน)สปอร์ทนต่อกรดดังนั้นจึงย้อมเป็นสีแดงตามวิธี Aujeszky หรือตามวิธี Ziehl-Neelsen และเซลล์พืชเป็นสีน้ำเงิน
รูปร่างของข้อพิพาทอาจเป็นวงรีทรงกลม ตำแหน่งในเซลล์คือเทอร์มินัลเช่น ที่ส่วนท้ายของแท่ง (ในสาเหตุของบาดทะยัก), subterminal - ใกล้กับส่วนท้ายของแท่ง (ในเชื้อโรคของโรคโบทูลิซึม, โรคเนื้อตายเน่าของก๊าซ) และส่วนกลาง (ในแบคทีเรียแอนแทรกซ์) สปอร์ยังคงอยู่เป็นเวลานานเนื่องจากมีเปลือกหลายชั้น แคลเซียมไดปิโคลิเนต ปริมาณน้ำต่ำ และกระบวนการเผาผลาญที่ซบเซา ภายใต้สภาวะที่เอื้ออำนวย สปอร์จะงอกผ่านสามขั้นตอนต่อเนื่องกัน: การเปิดใช้งาน การเริ่มต้น การงอก
8. รูปแบบหลักของแบคทีเรีย
แบคทีเรียทั่วโลก (cocci)มักเป็นทรงกลม แต่อาจเป็นวงรีหรือถั่วเล็กน้อย Cocci สามารถอยู่ได้โดยลำพัง (micrococci); เป็นคู่ (diplococci); ในรูปแบบของลูกโซ่ (สเตรปโตคอคซี) หรือพวงองุ่น (สแตฟิโลคอคซี) แพ็คเกจ (ซาร์ซินาส) Streptococci สามารถทำให้เกิดต่อมทอนซิลอักเสบและไฟลามทุ่ง Staphylococci - กระบวนการอักเสบและเป็นหนองต่างๆ
แบคทีเรียรูปแท่งที่พบมากที่สุด. แท่งสามารถเป็นแบบเดี่ยว ต่อเป็นคู่ (ไดพโพลแบคทีเรีย) หรือเป็นโซ่ (สเตรปโตแบคทีเรีย) แบคทีเรียที่มีรูปร่างเป็นแท่ง ได้แก่ Escherichia coli เชื้อโรคของเชื้อ Salmonellosis โรคบิด ไข้ไทฟอยด์ วัณโรค ฯลฯ แบคทีเรียรูปแท่งบางชนิดสามารถก่อตัวได้ภายใต้สภาวะที่ไม่เอื้ออำนวย ข้อพิพาทแท่งสร้างสปอร์เรียกว่า แบคทีเรียแบคทีเรียรูปแกนเรียกว่า คลอสตริเดีย
การสร้างสปอร์เป็นกระบวนการที่ซับซ้อน สปอร์แตกต่างจากเซลล์แบคทีเรียปกติอย่างมาก พวกมันมีเปลือกหนาแน่นและมีน้ำเพียงเล็กน้อย พวกมันไม่ต้องการสารอาหารและการสืบพันธุ์จะหยุดลงอย่างสมบูรณ์ สปอร์สามารถทนต่อการทำให้แห้ง อุณหภูมิสูงและต่ำได้เป็นเวลานาน และสามารถอยู่ในสภาพที่ทำงานได้เป็นเวลาหลายสิบและหลายร้อยปี (สปอร์ของแอนแทรกซ์ โบทูลิซึม บาดทะยัก ฯลฯ) เมื่ออยู่ในสภาพแวดล้อมที่เอื้ออำนวยสปอร์จะงอกนั่นคือพวกมันกลายเป็นรูปแบบการขยายพันธุ์ตามปกติ
แบคทีเรียที่ซับซ้อนสามารถอยู่ในรูปแบบของเครื่องหมายจุลภาค - vibrios โดยมีหยิกหลายอัน - spirilla ในรูปแบบของแท่งเกลียวบาง ๆ - สไปโรเชต Vibrios เป็นสาเหตุของอหิวาตกโรคและสาเหตุของซิฟิลิสคือ spirochete
9. คุณสมบัติของสัณฐานวิทยาของ rickettsia และ chlamydia
Rickettsia เป็นจุลินทรีย์แกรมลบขนาดเล็กที่โดดเด่นด้วยความหลากหลายที่เด่นชัด - พวกมันก่อตัวเป็น cocci, รูปแท่งและรูปแบบใย (รูปที่ 22) ขนาด Rickettsia แตกต่างกันไปตั้งแต่ 0.5 ถึง 3-4 ไมครอนความยาวของรูปแบบใยถึง 10-40 ไมครอน พวกมันไม่ก่อตัวเป็นสปอร์และแคปซูล แต่มีสีแดงตาม Zdrodovsky
Chlamydia มีลักษณะเป็นทรงกลม รูปไข่ หรือรูปแท่ง ขนาดของพวกเขาผันผวนภายใน 0.2-1.5 ไมครอน สัณฐานวิทยาและขนาดของหนองในเทียมขึ้นอยู่กับระยะของวัฏจักรการพัฒนาภายในเซลล์ ซึ่งมีลักษณะเฉพาะโดยการเปลี่ยนแปลงของการก่อตัวเบื้องต้นทรงกลมขนาดเล็กให้กลายเป็นร่างกายเริ่มต้นขนาดใหญ่ที่มีการแบ่งไบนารี ก่อนที่จะแบ่งอนุภาค Chlamydia จะถูกห่อหุ้มในรูปแบบที่คล้ายกับแคปซูลแบคทีเรีย Chlamydia stain "* ตาม Romanovsky-Giemsa, แกรมลบ, มองเห็นได้ชัดเจนในการเตรียม intravital ด้วยกล้องจุลทรรศน์แบบเฟสคอนทราสต์
10. โครงสร้างและชีววิทยาของไมโคพลาสมา
Mycoplasmatales ลำดับเดียวเท่านั้นที่อยู่ในคลาส Mollicutes ตัวแทนของคำสั่งนี้ - mycoplasmas -
ต่างจากแบคทีเรียตรงที่ไม่มีผนังเซลล์ แต่มีเยื่อไซโตพลาสมิกไลโปโปรตีนสามชั้นแทน ขนาดของ mycoplasmas ผันผวนภายใน 125-250 ไมครอน มีลักษณะกลม วงรีหรือเส้นใยแกรมลบ
Mycoplasmas ทำซ้ำโดยการแบ่งแยกแบบไบนารี เช่นเดียวกับแบคทีเรียส่วนใหญ่ โดยเฉพาะอย่างยิ่งหลังจากการก่อตัวของ coccoid ขนาดเล็ก (ร่างกายเบื้องต้น, EB) ในโครงสร้างเส้นใย
Mycoplasmas สามารถแตกหน่อและแบ่งส่วนได้ หน่วยการสืบพันธุ์ขั้นต่ำคือ ET (0.7-0.2 ไมครอน) ส่วนประกอบหลักของเยื่อหุ้มเซลล์คือคอเลสเตอรอล Mycoplasmas ไม่สามารถสร้างคอเลสเตอรอลและใช้ประโยชน์ได้จากเนื้อเยื่อหรือสารอาหารที่เสริมด้วยการแนะนำ การย้อมสีแกรมเป็นลบ แต่การย้อมแบบโรมานอฟสกี-เกียมซาให้ผลลัพธ์ที่ดีที่สุด มัยโคพลาสมาต้องการเงื่อนไขการเพาะปลูก: ต้องเติมซีรั่มพื้นเมือง, โคเลสเตอรอล, กรดนิวคลีอิก, คาร์โบไฮเดรต, วิตามินและเกลือต่าง ๆ ลงในอาหารเลี้ยงเชื้อ บนสื่อที่มีความหนาแน่นสูง พวกมันจะสร้างโคโลนีโปร่งแสงขนาดเล็กที่มีลักษณะเฉพาะโดยมีจุดศูนย์กลางเม็ดเล็กที่ยกขึ้น ทำให้พวกมันดูเหมือน "ไข่ทอด" ในสื่อที่มีเลือด มัยโคพลาสมาบางชนิดจะให้ a- และ beta-hemolysis ในสื่อกึ่งของเหลว มัยโคพลาสมาจะเติบโตตามแนวการฉีด ก่อตัวเป็นอาณานิคมที่กระจัดกระจายและแตกเป็นเสี่ยงๆ ในตัวกลางที่เป็นของเหลวทำให้เกิดฝ้าหรือสีเหลือบเล็กน้อย บางสายพันธุ์สามารถสร้างฟิล์มมันเยิ้มที่บางที่สุดได้ ในมนุษย์ ตัวแทนของสกุล Mycoplasma, Ureaplasma และ Acholeplasma ถูกแยกออก ซึ่งรวมถึงสายพันธุ์ที่ทำให้เกิดโรคและ saprophytic