По расположению жгутиков бактерии делятся на. Органоиды движения: функции и строение, особенности движения простейших
Строение. Около половины известных видов бактерий на поверхности имеют органы движения - волнообразно изогнутые жгутики. Масса жгутиков составляет до 2 % сухой массы бактерии. Длина жгутика больше длины тела микроорганизма и составляет 3–12 мкм; толщина жгутика 0,02 мкм, причем полярные жгутики более толстые, чем перитрихиальные.
Жгутики состоят из белка флагеллина (лат. flagella - жгутик), который по своей структуре относится к сократительным белкам типа миозина. В составе жгутика имеется либо одна гомогенная белковая нить, либо 2–3 нити, плотно свернутые в косу. Нить жгутика - жесткая спираль, закрученная против часовой стрелки; шаг спирали специфичен для каждого вида бактерий.
Число, размеры и расположение жгутиков являются признаками, постоянными для определенного вида, и учитываются при систематике. Однако у некоторых бактерий могут образовываться жгутики разных типов. Кроме того, наличие жгутиков зависит от условий внешней среды: на твердых средах при длительном культивировании бактерии могут утратить жгутики, а на жидких - вновь приобрести. Количество и расположение жгутиков у одного и того же вида может определяться стадией жизненного цикла. Следовательно, не стоит переоценивать таксономическое значение этого признака.
Классификация бактерий по числу и расположению жгутиков:
1. Атрихи - жгутики отсутствуют.
2. Монотрихи - один жгутик, расположенный на одном из полюсов клетки (род Vibrio) - монополярное монотрихальное расположение жгутиков, самые подвижные бактерии.
3. Политрихи - много жгутиков:
– лофотрихи - пучок жгутиков на одном полюсе клетки (роды Pseudomonas , Burkholderia ) - монополярное политрихальное расположение жгутиков;
– амфитрихи - на каждом полюсе клетки расположено по пучку жгутику (род Spirillum) - биполярное политрихальное расположение жгутиков;
– перитрихи - жгутики расположены без определенного порядка по всей поверхности клетки (сем. Enterobacteriaceae (роды Escherichia, Proteus ), сем. Bacillaceae, сем. Clostidiaceae ), число жгутиков от 6 до 1000 на клетку в зависимости от вида бактерий (рис. 7).
Рис.7. Варианты расположения жгутиков у бактерий:
1 - монотрих, 2 - лофотрих;
3 - амфитрих; 4 - перитрих.
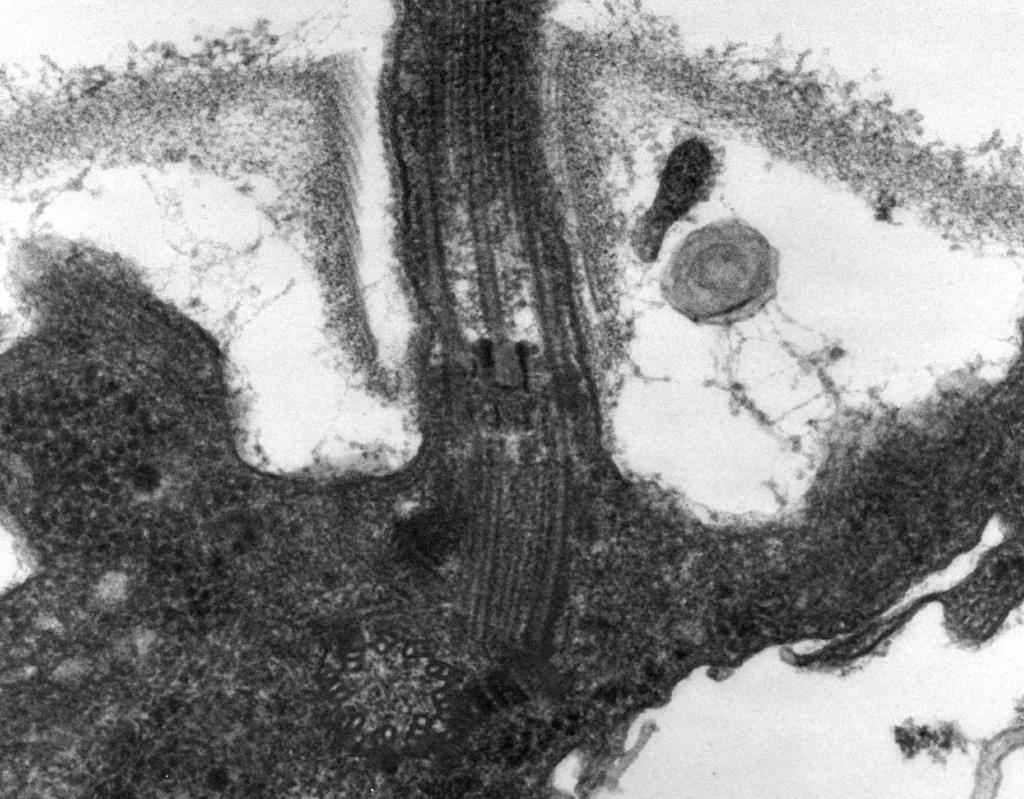
– При электронной микроскопии обнаружено, что жгутик состоит из трех частей: спиральной нити, крюка и базального тела (рис. 8).
Основную часть жгутика составляет длинная спиральная нить (фибрилла) - жесткий полый цилиндр диаметром около 120 нм, состоящий из белка флагеллина. По длине нити белковые молекулы образуют 11 рядов и уложены в виде спирали. В процессе роста нити белковые молекулы, синтезированные внутри клетки, проходят через полость цилиндра и пристраиваются в спираль на ее конце. На конце жгутика имеется белковая шапочка (крышечка), закрывающая отверстие цилиндра и препятствующая выходу молекул белка в окружающую среду. Длина нити жгутика может достигать нескольких микрометров. У некоторых видов бактерий жгутик снаружи дополнительно покрыт чехлом. У поверхности КС спиральная нить переходит в утолщенную изогнутую структуру - крюк.

Рис. 8. Схема строения жгутика
2. Крюк (толщиной 20–45 нм) вблизи поверхности клетки - относительно короткий цилиндр, состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом.
3. Базальное тело находится в основании жгутика и обеспечивает его вращение. Базальное тело содержит 9–12 различных белков и состоит из двух или четырех дисков (колец), нанизанных на стержень, являющийся продолжением крюка. Эти кольца вмонтированы в ЦПМ и КС. Два внутренних кольца (M и S) - обязательные составные части базального тела. M-кольцо локализовано в ЦПМ, S-кольцо располагается в периплазматическом пространстве грамотрицательных или в пептидогликановом мешке грамположительных бактерий. Два наружных кольца (D и L) необязательны для движения, так как имеются только у грамотрицательных бактерий, локализованы соответственно в пептидогликановом слое и в наружной мембране КС. Кольца S, D и L неподвижны и служат для фиксации жгутика в КС. Вращение жгутика определяется вращением M–кольца, встроенного в ЦПМ клетки. Таким образом, особенности строения базального тела жгутика определяются строением КС.
Функционально базальное тело представляет собой электромотор, работающий на протонах. М–кольцо базального тела (вращающийся ротор), окружено мембранными белками, имеющими отрицательные заряды (статор мотора). Бактериальная клетка обладает эффективным механизмом, позволяющим превращать электрохимическую энергию в механическую. Поэтому на работу жгутика бактерия тратит около 0,1 % всей расходуемой ею энергии. При работе жгутика используется протондвижущая сила, которая обеспечивается разностью концентраций протонов на внешней и внутренней сторонах мембраны (на внешней стороне их больше) и наличием более отрицательного заряда на внутренней стороне мембраны. Протондвижущая сила заставляет протоны проходить через базальное тело внутрь клетки, при этом они задерживаются на определенных участках ротора, придавая им положительный заряд, затем протоны уходят внутрь клетки. Заряженные участки расположены таким образом, что возникает сила притяжения между заряженными участками ротора и статора, М-кольцо начинает вращаться со скоростью около 300 об/c. Механизм вращения: зарядка–перезарядка группы COOH в аминокислотах. Для полного оборота кольца через базальное тело должно пройти 500–1000 протонов. Вращение М-кольца через жестко связанную с ним ось и крюк передается нити жгутика, которая функционирует как пропеллер или корабельный винт. Бактерия плывет до тех пор, пока работает винт, вклад инерции исключительно мал.
Кроме того, бактерии, даже мертвые, находящиеся в водной среде, перемещаются в результате броуновского движения. Бактериальная клетка все время подвержена ударам окружающих молекул, находящихся в тепловом движении. Удары, наносимые с разных сторон, бросают бактерию из стороны в сторону.
Тип движения жгутиков - вращательный. Существуют два вида движения: прямолинейное и кувыркание (периодические случайные изменения направления движения). Когда жгутики вращаются против хода часовой стрелки (около 1 секунды), с частотой 40–60 об./с (близко к скорости среднего электромотора), их нити сплетаются в единый жгут (рис. 9а). Вращение жгутиков передается клетке. Так как клетка намного массивнее жгутика, она начинает двигаться по прямой в противоположном направлении, со скоростью в 3 раза меньшей, чем скорость движения жгутика.
Так обеспечивается поступательное движение клетки, скорость которого в жидкой среде для разных видов бактерий составляет 20–200 мкм/с (это соответствует примерно 300–3000 длин тела в минуту) и более медленное перемещение по поверхности твердых сред.
Плыть целенаправленно в одном направлении бактерия может не более 3 с, затем удары окружающих молекул разворачивают ее в случайном направлении. Поэтому бактерии выработали механизм спонтанного изменения направления движения - переключение жгутикового мотора. Когда он начинает вращаться по ходу часовой стрелки (около 0,1 с), бактерия останавливается и переворачивается (совершает «кувырок») в случайном направлении. При этом жгутики разлетаются в разные стороны (рис. 9б). У амфитрихов при движении один пучок жгутиков вывернут наизнанку (подобно вывернутому ветром зонту). Затем мотор снова переходит к вращению против часовой стрелки, и бактерия опять плывет по прямой, но уже в другом, случайном направлении.
Жгутики могут также изменять направление движения в ответ на внешний стимул. Если бактерия перемещается в сторону оптимальной концентрации аттрактанта, жгутики проталкивают клетку через среду, ее прямолинейное движение становится более длительным, а частота кувырканий более низкой, что позволяет ей в конечном итоге перемещаться в нужном направлении.
Известны случаи существования бездействующих (парализованных) жгутиков. Для движения жгутиковых бактерий необходима интактность (неповрежденность) КС. Обработка клеток лизоцимом, приводящая к удалению пептидогликанового слоя КС, вызывает потерю способности бактерий к движению, хотя жгутики остаются при этом неповрежденными.
Таксисы бактерий. Пока окружающая среда остается неизменной, бактерии плавают беспорядочно. Однако совершенно однородной окружающая среда бывает редко. Если среда неоднородна, бактерии проявляют элементарные поведенческие реакции: активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие генетически детерминированные целенаправленные перемещения бактерий называют таксисами. В зависимости от фактора различают хемотаксис (частный случай - аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис.
Хемотаксис - движение в определенном направлении относительно источника химического вещества. Химические вещества делят на две группы: инертные и вызывающие таксисы - хемоэффекторы. Среди хемоэффекторов есть вещества, привлекающие бактерий, - аттрактанты (сахара, аминокислоты, витамины, нуклеотиды), и вещества, их отпугивающие, - репелленты (некоторые аминокислоты, спирты, фенолы, неорганические ионы). Аттрактантом для аэробных и репеллентом для анаэробных прокариот является молекулярный кислород. Аттрактанты часто представлены пищевыми субстратами, хотя не все вещества, необходимые для организма, выступают в качестве аттрактантов. Также не все ядовитые вещества служат репеллентами и не все репелленты вредны. Таким образом, бактерии способны реагировать не на любые соединения, а только на определенные и различные для разных бактерий.
В поверхностных структурах бактериальной клетки есть специальные белковые молекулы - рецепторы, специфически соединяющиеся с определенным хемоэффектором, при этом молекула хемоэффектора не изменяется, а в молекуле рецептора происходят конформационные изменения. Рецепторы расположены неравномерно по всей поверхности клетки, а сконцентрированы на одном из полюсов. Состояние рецептора отражает внеклеточную концентрацию соответствующего эффектора.
Хемотаксис имеет приспособительное значение. Например, формы холерного вибриона с нарушенным хемотаксисом оказываются менее вирулентными.
Аэротаксис - нуждающиеся в молекулярном кислороде бактерии скапливаются вокруг пузырьков воздуха, попавших под покровное стекло.
Фототаксис - движение к свету или от него, свойствен фототрофным бактериям, использующим свет в качестве источника энергии.
Магнитотаксис - способность водных бактерий, содержащих кристаллики железосодержащих минералов, плыть вдоль линий магнитного поля Земли.
Термотаксис - движение в сторону изменения температуры, что имеет большое значение для некоторых патогенных бактерий.
Вискозитаксис - способность реагировать на изменение вязкости раствора. Обычно бактерии стремятся в среду с большей вязкостью, что имеет большое значение для патогенных видов.
Скольжение бактерий. Способность к скольжению с небольшой скоростью (2–11 мкм/с) по твердому или вязкому субстрату обнаружена у некоторых прокариот, напр., микоплазм.
Существуют несколько гипотез, объясняющих скользящее движение. Согласно гипотезе реактивного движения оно обусловлено выделением слизи через многочисленные слизевые поры в КС, в результате чего клетка отталкивается от субстрата в направлении, противоположном направлению выделения слизи. Согласно гипотезе «бегущей волны» скользящее движение у подвижных безжгутиковых форм связано с наличием между пептидогликановым слоем и наружной мембраной КС тонкого белкового слоя из упорядоченно расположенных фибрилл, аналогичных нитям жгутиков. Вращательное движение фибрилл, «запускаемое» этими структурами, приводит к появлению на поверхности клетки «бегущей волны» (движущихся микроскопических выпуклостей КС), в результате чего клетка отталкивается от субстрата. Наконец, у некоторых скользящих бактерий описаны структуры, напоминающие базальные тела жгутиковых форм.
Функции жгутиков:
1. Обеспечивают адгезию - начальную стадию инфекционного процесса.
2. Обеспечивают подвижность бактерий.
3. Определяют антигенную специфичность, это Н-антиген.
Выявление жгутиков:
1. Фазовоконтрастная микроскопия нативных препаратов («раздавленной» и «висячей» капли). Микроскопически подвижность определяют у клеток суточной культуры. Для того чтобы отличить подвижность от пассивного броуновского движения, к капле исследуемой культуры добавляют каплю 5 %–ного водного раствора фенола, активное движение в этом случае прекращается.
2. Темнопольная микроскопия нативных препаратов.
3. Световая микроскопия окрашенных красителями или металлами препаратов. Так как жгутики очень легко повреждаются при приготовлении препарата, в повседневной практике эти методы используется редко.
Для окраски жгутиков используют клетки, выращенные на скошенном агаре. Бактериальной петлей отбирают клетки, находящиеся у конденсационной воды и осторожно переносят в стерильную дистиллированную воду такой же температуры, что и температура инкубирования бактерий на скошенном агаре, а бактерии с петли не стряхивают, а осторожно погружают в воду. Пробирку с бактериями оставляют при комнатной температуре на 30 мин. Используют химически чистое (вымытое в хромовой смеси) стекло, на которое наносят 2–3 капли суспензии. Суспензию распределяют по поверхности стекла, осторожно его наклоняя. Высушивают препарат на воздухе.
Жгутики очень тонкие, поэтому их можно обнаружить только при специальной обработке. Вначале при помощи протравки достигается разбухание и увеличение их размера, а затем производится окраска препарата, благодаря чему они становятся видимыми при световой микроскопии.
Чаще используют метод серебрения по Морозову (рис. 10):
– препарат фиксируют раствором ледяной уксусной кислоты 1 минуту, промывают водой;
– наносят раствор таннина (дубящий, делающий жгутики более плотными) на 1 мин, промывают водой;
– обрабатывают препарат при подогревании импрегнирующим раствором азотнокислого серебра 1–2 мин, промывают водой, высушивают и микроскопируют.
При микроскопии видны темно-коричневые клетки и более светлые жгутики.

Рис. 10. Выявление жгутиков методом серебрения
Рис. 11. Выявление жгутиков
методом электронной микроскопии
4. Электронная микроскопия препаратов, напыленных тяжелыми металлами (рис. 11).
5. Косвенно - по характеру роста бактерий при посеве в полужидкий 0,3 %–ный агар. После инкубирования посевов в термостате в течение 1–2 сут отмечают характер роста бактерий:
– у неподвижных бактерий (напр., S. saprophyticus ) наблюдается рост по ходу укола - «гвоздь», а среда прозрачна;
– у подвижных бактерий (напр., Е. со1i ) наблюдается рост в стороны от укола, по всему столбику агара - «елочка», и диффузное помутнение среды.
Все бактерии подразделяются на подвижные и неподвижные. Органами движения у бактерий являются жгутики. Они состоят из белка флагеллина, который по своей структуре относится к сократительным белками типа миозина.
Основанием жгутика
является базальное тельце, состоящее из системы дисков (блефаропласта: 1 диск - наружная сторона клеточной стенки, 2 диск - внутренняя сторона клеточной стенки, 3 диск - цитоплазматическая мембрана), "вмонтированных" в цитоплазматическую мембрану и клеточную стенку. Длина жгутика больше длины тела самого микроба.
По числу жгутиков
и их расположению подвижные микроорганизмы подразделяются на:
1. Монотрихи, имеющие на конце тела один жгутик (самые подвижные). Например, Vibrio cholerae.
2. Лофотрихи, имеющие пучок жгутиков на одном из полюсов клетки. Например, Burkholderia (Pseudomonas) pseudomalei - возбудитель мелиоидоза.
3. Амфитрихи, имеющие жгутик на обоих полюсах клетки. Например, Spirillum volutans.
4. Перитрихи, имеющие жгутики по всему периметру клетки. Например, Escherichia coli, Salmonella typhi.
Выявление жгутиков. Жгутики очень тонкие, поэтому их можно обнаружить только при специальной обработке. В частности, вначале при помощи протравы достигается разбухание и увеличение их размера, а затем производится окраска препарата, благодаря чему они становятся видимыми при световой микроскопии. Выявлять жгутики можно окраской по Морозову, Леффлеру, а также электронной микроскопией. Обнаружить жгутики можно и по активной подвижности бактерий.
Движение микробов наблюдают в препаратах "раздавленной" и "висячей" капли из живых культур. Микроскопируют эти препараты сухим или иммерсионным объективом в темном поле или в фазовом контрасте. Кроме того, подвижность можно определить по характеру роста бактерий в полужидком агаре.
Пили у бактерий.
Пили (pili), синонимы: ворсинки, фимбрии, - тонкие полые нити белковой природы, покрывающие поверхность бактериальных клеток. В отличие от жгутиков не выполняют двигательную функцию.
Пили отходят от поверхности клетки и состоят из белка пилина .
По своему функциональному назначению подразделяются на 2 типа.
1) Пили первого типа имеются у большинства бактерий, поэтому они получили название "ворсинки общего типа" (common pili). Обусловливают прикрепление или адгезию бактерий к определенным клеткам организма хозяина. Адгезия является первоначальной стадией любого инфекционного процесса.
2) Пили второго типа (синонимы: конъюгативные, или половые - sex pili) имеются только у бактерий-доноров, имеющих специальную плазмиду. Их количество невелико - 1-4 на клетку.
Половые пили выполняют следующие функции:
1. Участвуют в передаче генетического материала от одной клетки к другой при конъюгации бактерий.
2. На них адсорбируются специфические вирусы бактерий – бактериофаги
Споры бактерий, условия образования, расположение, механизм и этапы окраски по Ауески.
Споры - своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки.
Спорообразование - это способ сохранения вида (генофора) во внешней среде при неблагоприятных условиях, а не способ размножения.
Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). Внутри бактериальной клетки образуется одна спора (эндоспора).
Стадии спорообразования
1. Подготовительная. В цитоплазме бактерий образуется уплотненный участок, не имеющий свободной воды, называемый "спорогенной зоной", в которой содержится нуклеоид.
2. Стадия предспоры (проспоры). Вокруг спорогенной зоны образуется оболочка из двойной цитоплазматической мембраны.
3. Образование кортекса, состоящего из пептидогликана и наружной мембраны с повышенным содержанием солей кальция и липидов.
4. Стадия созревания. С внешней стороны наружной мембраны образуется оболочка споры, после чего вегетативная часть клетки лизируется, освобождая спору.
Развитие микробиологии принесло в последние десятилетия множество открытий. И одно из них - это особенности движения жгутиковых бактерий. Устройство двигателей этих древнейших организмов оказалось очень сложным и по принципу своей работы очень отличающимся от жгутиков наших ближайших эукариотических родственников простейших. Вокруг двигателя жгутиковой бактерии разгорелись самые жаркие споры между креационистами и эволюционистами. О бактериях, их жгутиковых моторах и о многом другом - данная статья.
Общая биология
Для начала вспомним, что это за организмы и какое место они занимают в системе органического мира на нашей планете. Домен Bacteria объединяет огромное количество одноклеточных прокариотических (без оформленного ядра) организмов.
Эти живые клетки появились на арене жизни почти 4 миллиарда лет назад и были первыми поселенцами планеты. Они могут быть самой разной формы (кокки, палочки, вибрионы, спирохеты), но большинство из них - жгутиковые.
Где живут бактерии? Везде. На планете их обитает более 5×10 30 . В 1 грамме почвы их насчитывается порядка 40 миллионов, в нашем организме обитает до 39 триллионов. Их можно встретить на дне Марианской впадины, в горячих «черных курильщиках» на дне океанов, во льдах Антарктиды, а на ваших руках в данный момент находится до 10 миллионов бактерий.
Значение неоспоримо
Несмотря на микроскопические размеры (0,5-5 мкм) их общая биомасса на Земле больше, чем биомасса животных и растений, вместе взятых. Их роль в круговороте веществ незаменима, а их свойства консументов (разрушителей органики) не позволяют планете покрыться горами трупов.
Ну и не стоит забывать о патогенах: возбудителями чумы, оспы, сифилиса, туберкулеза и множества других инфекционных заболеваний тоже являются бактерии.
Бактерии нашли применение и в хозяйственной деятельности человека. Начиная от пищевой промышленности (кисломолочные продукты, сыры, квашеные овощи, алкогольные напитки), «зеленой» экономике (биотопливо и биогаз) до методов клеточной инженерии и производства лекарственных препаратов (вакцины, сыворотки, гормоны, витамины).

Общая морфология
Как уже говорилось, у этих одноклеточных представителей жизни нет ядра, их наследственный материал (молекулы ДНК в виде кольца) расположены в определенном участке цитоплазмы (нуклеоид). Клетка их имеет плазматическую мембрану и плотную капсулу, образованную пептидогликаном муреином. Из клеточных органелл у бактерий есть митохондрии, могут быть хлоропласты и другие структуры с различными функциями.
Большинство бактерий - жгутиковые. Плотная капсула на поверхности клетки не позволяет им передвигаться при помощи изменения самой клетки, как это делают амебы. Их жгутики - это плотные белковые образования различной длины и диаметром около 20 нм. Одни бактерии имеют единственный жгутик (монотрихи), у других их два (амфитрихи). Иногда жгутики расположены пучками (лофотрихи) или же покрывают всю поверхность клетки (перитрихи).
Многие из них живут в виде одиночных клеток, но некоторые образуют скопления (пары, цепочки, филаменты, гифы).

Особенности движения
Жгутиковые бактерии могут двигаться по-разному. Одни движутся только вперед, а направление меняют путем кувыркания. Некоторые способны к подергиваниям, другие передвигаются путем скольжения.
Жгутики бактерий выполняют функции не только клеточного «весла», но могут быть и «абордажным» орудием.
Совсем до недавнего времени считали, что жгутик бактерии виляет как хвост у змеи. Последние исследования показали, что жгутик бактерий устроен куда сложнее. Он работает как турбина. Присоединенный к приводу, он вращается в одном направлении. Привод, или жгутиковый мотор бактерии, - это сложная молекулярная структура, которая работает как мышца. С тем отличием, что мышца после совершения сжатия должна расслабляться, а мотор бактерии работает постоянно.

Наномеханизм работы жгутика
Не углубляясь в биохимию движения, отметим, что в создании привода жгутика учувствует до 240 белков, которые делятся на 50 молекулярных составляющих с определенной функцией в системе.
В этой двигательной системе бактерии есть ротор, который движется, и статор, который обеспечивает это движение. Есть приводной вал, втулка, сцепление, тормоза и ускорители
Этот миниатюрный двигатель позволяет бактерии совершать перемещения на расстояния в 35 раз большие, чем ее собственный размер всего лишь за 1 секунду. При этом на работу самого жгутика, совершающего 60 тысяч оборотов в минуту, организм тратит всего 0,1 % всей энергии, которую расходует клетка.
Удивительно еще и то, что все запчасти своего двигательного механизма бактерия может заменять и ремонтировать «на ходу». Просто представьте, что вы летите в самолете. А техники меняют лопасти работающего мотора.
Жгутиковая бактерия против Дарвина
Двигатель, способный работать со скоростью до 60 000 оборотов в минуту, самочинящийся и использующий в виде топлива всего лишь углеводы (сахара), имеющий устройство сродни электрическому двигателю - могло ли такое устройство возникнуть в процессе эволюции?
Именно этот вопрос задал себе в 1988 году доктор биологических наук Майкл Бихи. Он ввел в биологию понятие несократимой системы - системы, в которой все ее части одновременно необходимы для обеспечения ее работы, а удаление хоть одной части приводит к полному нарушению ее функционирования.
С позиции эволюции Дарвина, все структурные изменения в организме происходят постепенно и отбираются естественным отбором только успешные.
Выводы М. Бихи, изложенные в книге «Черный ящик Дарвина» (1996): двигатель жгутиковой бактерии - неделимая система из более чем 40 частей, и отсутствие хоть одной приведет к полной нефункциональности системы, а значит, эта система не могла произойти путем естественного отбора.

Бальзам для креационистов
Теория сотворения в изложении ученого и профессора биологии, декана факультета биологических наук в Лихайском университете Бетлехема (США) М. Бихи сразу привлекла внимание служителей церкви и сторонников теории божественного происхождения жизни.
В 2005 году в США даже состоялся судебный процесс, где Бихи выступал свидетелем со стороны сторонников теории «разумного дизайна», на котором рассматривался вопрос введения в школах Дувра изучения креационизма на курсе «О пандах и людях». Процесс был проигран, преподавание такого предмета признали противоречащим действующей конституции.
Но споры креационистов и эволюционистов продолжаются и сегодня.
Жгутик – это поверхностная структура бактериальной клетки, которая служит им для движения в жидких средах.
В зависимости от расположения жгутиков, бактерии делятся на (Рис.1):
Перитрихиальные
Смешанные
Полюсные
Подполюсные
Полюсные жгутики – один или более жгутиков расположены на одном (монополярно) или обоих (биполярно) полюсах клетки и основание параллельно длинной оси клетки.
Подполюсные жгутики (субполярные) – один или более жгутиков расположены в месте перехода боковой поверхности в полюс клетки на одном или двух ее концах. В основании – прямой угол с длинной осью клетки.
Боковые жгутики (латеральные) - один или более жгутиков в виде пучка расположены в средней точке одной из половин клетки.
Перитрихиальные жгутики – расположены по всей поверхности клетки по одному или пучками, полюса обычно их лишены.
Смешанные жгутики – два или несколько жгутиков расположены в разных точках клетки.
В зависимости от числа жгутиков, различают:
Монотрихи – один жгутик
Политрихи – пучок жгутиков
Так же выделяют:
Лофотрихи – моноплярное политрихиальное расположение жгутиков.
Амфитрихи – биполярное политрихиальное расположение жгутиков.
Строение бактериального жгутика и базального тела. Жгутик.
Сам по себе жгутик устроен довольно просто: филамент, который крепится к базальному телу. Иногда между базальным телом и филаментом может быть вставлен изогнутый участок трубки, так называемый крюк, он толще филамента, участвует в гибком присоединении филамента к базальному телу.
По химическому составу жгутик на 98% состоит из белка флагеллина (flagellum – жгутик), он содержит 16 аминокислот, преобладают глутаминовая и аспарагиновая, незначительное количество ароматических аминокислоты отсутствуют триптофан, цистеин и цистин. Флагеллин облагает антигенной специфичностью, его называют Н-антиген. Жгутики бактерий не обладают АТФазной активностью.
Толщина жгутика 10 – 12 нм, длина 3-15 мкм.
Представляет собой жесткую спираль, закрученную против часовой стрелки. Вращение жгутика осуществляется также против часовой стрелки с частотой от 40 об/сек до 60 об/сек, что вызывает вращение клетки в противоположном направлении, но т.к. клетка намного тяжелее жгутика, то ее вращение медленнее от 12 до 14 об/сек.
Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом, который является продолжением клеточной стенки и, вероятно, имеет такое же строение.
Базальное тело
Базальное тело состоит из 4 деталей:
Стержень, стыкующийся с филаментом или с крюком
Два диска, нанизанных на стержень. (М и S)
Группа белковых комплексов (статоры)
Белковый колпачок
Бактерии, которые имеют внутреннюю и внешнюю мембрану имеют 2 дополнительных диска (P и L) и белковые структуры, которые находятся на внешней мембране вблизи базального тела, следовательно они не играют важной роли в движении.
Особенность строения базального тела определяется строением клеточной стенки: ее интактность необходима для движения жгутиков. Обработка клеток лизоцимом приводит к удалению пептидогликанового слоя с клеточной стенки, что ведет к потери движения, хотя строение жгутика нарушено не было.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
8. Основные формы бактерий
Шаровидные бактерии (кокки) имеют обычно форму шара, но могут быть немного овальной или бобовидной формы. Кокки могут располагаться поодиночке (микрококки); попарно (диплококки); в виде цепочек (стрептококки) или виноградных гроздьев (стафилококки), пакетом (сарцины). Стрептококки могут вызывать ангину и рожистое воспаление, стафилококки - различные воспалительные и гнойные процессы.
Палочковидные бактерии самые распространенные. Палочки могут быть одиночными, соединяться попарно (диплобактерии) или в цепочки (стрептобактерии). К палочковидным относятся кишечная палочка, возбудители сальмонеллеза, дизентерии, брюшного тифа, туберкулеза и др. Некоторые палочковидные бактерии обладают способностью при неблагоприятных условиях образовывать споры. Спорообразующие палочки называют бациллами. Бациллы, напоминающие по форме веретено, называют клостридиями.
Спорообразование представляет собой сложный процесс. Споры существенно отличаются от обычной бактериальной клетки. Они имеют плотную оболочку и очень малое количество воды, им не требуются питательные вещества, а размножение полностью прекращается. Споры способны длительно выдерживать высушивание, высокие и низкие температуры и могут находиться в жизнеспособном состоянии десятки и сотни лет (споры сибирской язвы, ботулизма, столбняка и др.). Попав в благоприятную среду, споры прорастают, т. е. превращаются в обычную вегетативную размножающуюся форму.
Извитые бактерии могут быть в виде запятой - вибрионы, с несколькими завитками - спириллы, в виде тонкой извитой палочки - спирохеты. К вибрионам относится возбудитель холеры, а возбудитель сифилиса - спирохета.
9. Особенности морфологии риккетсий и хламидий
Риккетсии представляют собой мелкие грамотрицательные микроорганизмы, характеризующиеся выраженным полиморфизмом-образуют кокковидные, палочковидные и нитевидные формы (рис. 22). Размеры риккетсии варьируют от 0,5 до 3-4 мкм, длина нитевидных форм достигает 10-40 мкм. Спор и капсул не образуют, окрашиваются по Здродовскому в красный цвет.
Хламидии имеют шаровидную, овоидную или палочковидную форму. Их размеры колеблются в пределах 0,2-1,5 мкм. Морфология и размеры хламидии зависят от стадии их внутриклеточного цикла развития, для которого характерно превращение небольшого шаровидного элементарного образования в крупное инициальное тельце с бинарным делением. Перед делением частицы хламидии обволакиваются образованием, напоминающим бактериальную капсулу. Хламидии окрашиваются"* по Романовскому-Гимзе, грамотрицательны, хорошо видны в прижизненных препаратах при фазово-контрастной микроскопии.
10. Строение и биология микоплазм.
К классу Mollicutes относится только один порядок Mycoplasmatales. Представители этого порядка - микоплазмы-
отличаются от бактерий отсутствием клеточной стенки. Вместо нее они содержат трехслойную липопротеидную цитоплазматическую мембрану. Размеры микоплазм колеблются в пределах 125-250 мкм. Они имеют форму круглых, овальных или нитевидных образований, грамотрицательны
Микоплазмы размножаются бинарным делением, подобно большинству бактерий, особенно после образования мелких кокковидных образований (элементарные тельца, ЭТ) в нитевидных структурах.
Микоплазмы способны к почкованию и сегментации. Минимальной репродуцирующейся единицей считают ЭТ (0,7-0,2 мкм). Основной компонент клеточной мембраны - холестерин. Микоплазмы не способны к образованию холестерина и утилизируют его из тканей или питательных сред, дополненных их внесением. По Граму окрашиваются отрицательно, но лучшие результаты даёт окраска по Романовскому-Гимзе. Микоплазмы требовательны к условиям культивирования: в питательные среды необходимо вносить нативную сыворотку, холестерин, нуклеиновые кислоты, углеводы, витамины и различные соли. На плотных средах образуют характерные мелкие полупрозрачные колонии с приподнятым зернистым центром, придающим им вид «яичницы-глазуньи». На средах с кровью некоторые виды микоплазм дают а- и бета-гемолиз. В полужидких средах микоплазмы растут по линии укола, формируя дисперсные, крош-коватые колонии. В жидких средах приводят к незначительному помутнению или опалесценции; некоторые штаммы способны образовывать тончайшую жирную плёнку. У человека выделяют представителей родов Mycoplasma, Ureaplasma и Acholeplasma, включающих патогенные и сапрофитические виды